优良睡眠、高效学习、美好生活

本文是 Piotr Wozniak 文章 Good sleep, good learning, good life 的中文译本,原文最初发表于 2012 年 5 月,2017 年 10 月 21 日更新。译文保留了原文章节结构,并将引用的图片本地化为本站静态资源。
Good sleep, good learning, good life
Dr Piotr Wozniak,2012 年 5 月(更新于 2017 年 10 月 21 日)
多年来,由于我在记忆和学习领域的专业投入,一直对睡眠研究保持着浓厚的兴趣。本文试图综合目前关于睡眠的已知知识,以期用于实际应用,尤其是那些需要高质量睡眠来支持学习或创造性成就的人。睡眠神经生理学是一门爆炸式增长的学科。许多目前存在争议的理论很快就会因新发现而被遗忘。因此,本文很可能很快就会过时(可对比 2000 年的旧版这里)。不过,关于睡眠的一些基本事实已是确立的共识,也能得出有益于人类创造力和智力成就的实际结论。本文中,我提供了一些指向研究论文和科普文章的链接,它们主张截然不同乃至相互矛盾的理论。请查阅其他来源,以确保你不会得到片面的观点!本文包含一些关于如何利用自由睡眠治疗失眠、提前和延迟相位移动综合征以及其他一些睡眠障碍的指引。如果你自己的经验能对本文提出的观点有所贡献,我将很乐意倾听(尤其是在学习和创造力的语境下)。
好的教育者坚持让孩子进行运动、娱乐和充足的睡眠:“这是大自然最伟大的强心剂。”
—— 约翰·洛克(John Locke)(1632-1704)
前言(Foreword)
每天醒来时神清气爽、心情愉悦、精神饱满地投入新一天,是每个人的梦想。遗憾的是,在现代社会,只有极少数人能真正过上这样的生活。然而,只要具备:
- 一点知识,以及
- 愿意在生活方式上做出一些牺牲,
这个梦想对大多数健康人来说都触手可及。
我希望这篇文章能汇集获得清爽睡眠所需的全部基础知识。至于”牺牲”,首先要明白一点:鱼与熊掌不可兼得。健康的睡眠可能与某些现代习惯、某些欲望或某些生活方式选择格格不入。在最坏的情况下,清爽的睡眠甚至可能与你的工作或长远目标相冲突。鉴于这一点,本文无法为每个人提供解决方案。此外,即便拥有充足的知识并把全部注意力都放在好睡眠上,要做到每天都有一个愉快、清醒的头脑也是一件困难的事。但是,我要郑重地说:对大多数人而言,在大多数夜晚拥有好睡眠是完全可行的!
这篇文章最初写于十年前。我长期以来一直关注记忆、学习和睡眠。在我的工作中,睡眠和氧气一样重要。随着我们都更深地迈入信息时代(Information Age)和知识经济(Knowledge Economy),这里讨论的问题对每个人都将变得越来越重要。在写下原文之后,我有幸接触到数百位饱受各种睡眠问题困扰的人,亲眼看到睡眠知识如何帮助解决他们的问题,也看到工业化时代如何为人们追求好睡眠和高生产力设置障碍。我目睹了一场真正的睡眠相位障碍(sleep phase disorders)流行病,见证了对多相睡眠(polyphasic sleep)兴趣的爆发,以及围绕睡眠问题整体兴趣的指数级增长。尽管我反复呼吁,许多人仍无法避免使用闹钟、考前通宵、把孩子从睡梦中拽起来去上学、睡前吞药片、让婴儿独自在婴儿床里哭到睡着(cry it out)等等。要不是因为”知识带来希望”这个事实,这幅图景会相当黯淡而令人警觉。只要有一点决心,每个人都能改善自己或孩子的睡眠。
这篇文章是关于睡眠生物学最重要、最有趣内容的汇编。它旨在帮助你掌握那些能让你获得高质量、令人神清气爽、从而提升心智能力所需的睡眠知识。文章解释了为什么睡眠对健康和大脑至关重要,论证了睡眠理应得到最高程度的尊重,而大多数人只要遵循既定规则,本可以获得出色的睡眠。
自最初撰写《Good sleep, good learning, good life》以来,睡眠科学取得了巨大进步。我自己使用 SleepChart 和 SuperMemo 等工具的工作,也为理解睡眠与学习之间的联系提供了一些有趣的线索。随着我在一些短文和常见问题解答(FAQ)中持续跟进睡眠科学的进展,supermemo.com 的一些访客抱怨说,有价值的信息碎片散落在站点各处,而没有以更百科全书式的方式组织在单篇文章里。于是有了这份综合汇编。我希望能保持对”有助于获得好睡眠的实用知识”的关注,同时仍想夹带一些对普通读者、以及对从事睡眠、记忆和学习研究的科学家可能有启发意义的较冷门研究发现。如果你认为我遗漏了任何他人应当知晓的重要内容,请让我知道。
由于文章已经长得离谱,你不妨从文末的总结开始读起。如果连总结都嫌长,这里是核心亮点:
- 敬畏睡眠,它是你获得高智商和好学习的工具。
- 自由睡眠(free running sleep) 能帮你解决许多睡眠问题。
- 双相睡眠(biphasic sleep) 的作息可能对创造型人群来说是最健康的作息。
- 不要为了上学叫醒孩子;如果他们无法按时醒来,让他们缺一两节课,或者考虑在家上学(homeschooling)。
- 让婴儿和幼儿按需睡眠,共同睡眠(co-sleeping)是个极好的主意(即便许多儿科医生会另有说法)。
- 运动、学习和睡眠 是你促进大脑生长的最佳工具!
- 避免用物质来调节睡眠与清醒,尤其是安眠药、酒精、违禁毒品、尼古丁和咖啡因。
关于注释
增量写作(Incremental writing):由于材料体量巨大,本文采用了一种叫做增量写作的技巧写成。增量写作有助于把大量早先的写作素材组织成一篇线性的文章。它的主要优点是,在快速处理来自不同来源的材料时,仍能保持合理的连贯性。用增量写作产出的文本特别适合借助增量阅读(incremental reading)来学习,因为它们会产生许多独立的小型维基百科式的子文章。但对线性读者而言,这可能意味着一定程度的冗长,以及令人烦恼的主题重复,对此我深表歉意。如果文章篇幅令人生畏,你可以尝试增量阅读(例如使用 SuperMemo 2004 免费版)?
参考文献:由于材料体量巨大,我无法为文中所有论断都提供出处。其中一些是常识,一些是普遍知识,另一些是我凭记忆或取自 SuperMemo 而未深挖到原始来源。如果你找不到某个具体论断的出处,请让我知道。
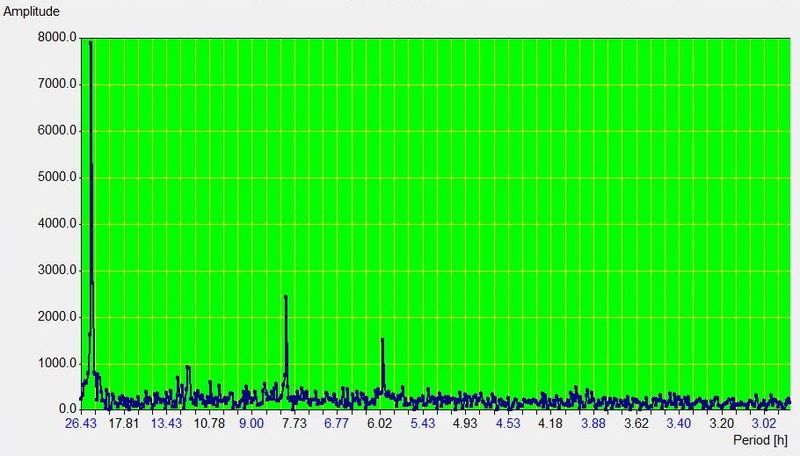
睡眠的重要性(Importance of sleep)
为什么理解睡眠如此重要?
太少人意识到睡眠有多重要!在现代社会绝大多数家庭里,闹钟是不可或缺的陈设。通过电灯、闹钟、安眠药和轮班工作,我们彻底破坏了睡眠这一过程。

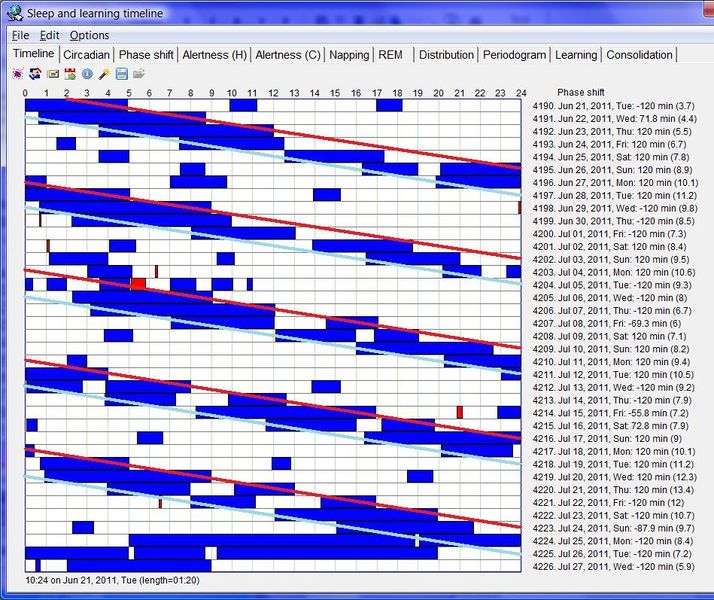
四份睡眠日志示例,说明现代人的睡眠模式千差万别,正如雪花各不相同。
在 20 世纪的一百年里,我们粗暴地干预了一个由数亿年演化精雕细琢、精细调节的过程。而我们直到最近才真正意识到,这种干预可能是阻碍工业化国家社会智力增长的最重要可预防因素之一!再过几年,我们看待闹钟和”睡眠调节”的方式,可能就像我们今天看待香烟、石棉材料或放射性化妆品这些”伟大”的人类发明一样。
对照下面的清单,看看哪些适用于你:
- 我经常在合适的时间难以入睡。
- 因为困倦,我经常觉得早上起床是一件极其痛苦的事。
- 我经常在学校或工作中感到昏昏欲睡。
- 与身体实际所需相比,我每天主动缩短 2-3 小时的睡眠。
- 我使用闹钟,并且非常讨厌它。
- 我每天喝大量的咖啡或可乐。
- 我经常在晚上进行 2-4 小时的长途小睡。
- 对我来说,上述至少有一项是经常性压力或生产力下降的来源。
我敢打赌,你有约 90% 的概率符合其中至少一项。也许这正是你读到这篇文章的原因。同样极有可能的是,你已经学会接受现状,认为自己对此无能为力。这篇文章或许能提示一些补救方法。然而坏消息是,要获得真正的解决方案,你很可能需要改变你的家庭生活、你的工作、你的老板,或者某些社会规则!
睡眠绝不仅仅是一种休息形式! 睡眠扮演着至关重要的生理功能,对你的智力发育是不可或缺的。那些不尊重自己睡眠的人,很难发挥出他们全部的智力潜能!
现代社会发展出了一套根深蒂固、极度无视睡眠的规则,而这在美国社会已经达到了病态的程度。以下是一些伤害睡眠的坏规则:
- 用闹钟把睡眠缩短是可以接受的。
- 轮班工作是可以接受的。
- 让人们在全球到处飞,而不太在意时差问题是可以接受的。
- 通过少睡多干来省时间是可以接受的。
- 为了按时上学把孩子从床上拽起来是可以接受的。
- 重要考试前熬夜是可以接受的,等等。
减少睡眠并不会让人立刻死亡(至少不会马上)。它确实让人感觉痛苦,但只要好好睡一晚就能恢复的那种轻松,似乎让睡眠看起来很廉价。甚至连吉尼斯世界纪录关于不眠的报道(Randy Gardner 1964 年长达 11 天的不眠挑战)都把不眠的影响淡化了。许多精神病学和心理学教科书至今仍声称,长时间不眠没有任何显著的副作用!这是错误的!吉尼斯世界纪录此后已因健康风险撤销了睡眠剥夺这一类别。
1992 年,比尔·克林顿在竞选总统时自豪地承认自己连续 48 小时没有睡觉,因为他真的很想成为下一任总统。前参议员鲍勃·多尔(Bob Dole)在 1996 年的竞选中”刷新”了这一纪录:我们已经连续工作了 78 个小时。我们还要干到 96 个小时。我们一直在为美国昼夜不停地奔走。 与多尔比肩的还有副总统阿尔·戈尔(Albert Gore Jr.),他在 2000 年 11 月 7 日选举日之前连续三天奔走竞选。选举之后,戈尔在与得克萨斯州州长乔治·W·布什那场胶着的选战中,仍通过反复”承认—撤回”的循环熬夜。而当巴拉卡·奥巴马在长达一年多的总统竞选后被问及最想要的圣诞礼物是什么时,他毫不犹豫地回答:8 小时的睡眠。
对睡眠不尊重的坏榜样,恰恰来自这个国家最重要的人!
然而,一些与睡眠剥夺相关的惊人事实已逐渐浮出水面。每年睡眠障碍给国家医疗带来 160 亿美元的额外成本(例如通过加剧高血压和心脏病)。这还不算工作中的事故和生产力损失。为此,国家睡眠障碍委员会(National Commission on Sleep Disorders)估计,睡眠剥夺每年因更高的压力和更低的工作场所生产力而造成 1500 亿美元的损失1。40% 的卡车事故可归因于疲劳和困倦,而午夜到早上 8 点之间单车商用卡车事故增加了 800%。一些重大工业灾难已被归因于睡眠剥夺(Mitler 等,19882)(包括三哩岛、切尔诺贝利、博帕尔毒气泄漏、Zeebrugge 海难以及 Exxon Valdez 油轮泄漏)。
自 20 世纪 20 年代以来,人们就知道睡眠能改善学习中的记忆保持。然而,直到千禧年之交,哈佛医学院精神病学副教授 Robert Stickgold 博士的研究才登上国际头条。Stickgold 博士的研究证明了一个早已为人所知却很少被重视的事实:睡眠对学习是必需的(Stickgold,20053)!睡眠不足时,我们对缩短睡眠前后所学事实的回忆能力都会下降。考前熬夜或许足以应付考试,但很少能在长期记忆中留下有用的痕迹。考试本身取代了知识,成了学习的主要目的!
减少睡眠,我们学得更少、发展得更慢、没那么聪明、做出更糟的决策、成就更少、生产力更低、更容易出错,并且损害了我们真正的智力潜能!
社会睡眠习惯的改变,将引发一场在学习、健康和生产力上规模之大鲜有人能想象的社会革命!“从历史来看,要出台保护社会免受因困倦而犯下灾难性错误之人危害的政策变革,似乎需要我们在看待睡眠的方式上进行根本性的转变。“(Merrill Mitler 博士)
我研究 SuperMemo 用户的学子性格已有二十多年。有几个因素决定了一个高效、持之以恒的优秀学子。以下是可能在学习上取得成功的人的一些特征:
- 高度乐观
- 睡眠良好
- 渴望知识
- 耐压
- 精力充沛,但能在学习时慢下来
以下是一些与有效学习能力相关性较差的不幸特征:
- 容易抑郁或情绪波动
- 睡眠有问题(尤其是失眠)
- 高压力水平
- 多动且注意力不集中
- 耐压能力低(吸烟者、滥用调节情绪物质者、酗酒者等)
睡眠良好,似乎是成功学习最重要的因素之一!
我们为什么睡眠?
多年来,睡眠的生理功能一直不清楚。在大多数人心中,睡眠与休息、与精神恢复的时间联系在一起。关于睡眠的恢复性、保护性和节能性理论一直相当流行,直到人们意识到:以抑制意识的方式来进行一次长时间的睡眠,似乎并不是演化处理资源耗竭、有毒废物或能量节约的正确方式。例如,肌肉并不需要完全关闭才能得到休息。睡眠的关键功能在以下实验中得到戏剧性的展示:长期被剥夺睡眠的大鼠通常在 2.5 周内死亡(详见后文:If you do not sleep, you die!)。
从演化角度看,睡眠是一个非常古老的现象,它显然扮演着对生存至关重要的角色。直到最近才被确切证明:睡眠的功能与学习相关(并非所有科学家都同意)!
研究人员早就知道海马体(hippocampus)——一个小的脑器官——对记忆形成的重要性。但海马体究竟有何特别之处,能把它与大脑皮层其他同样具有突触可塑性(synaptic plasticity,即存储记忆能力)的区域区分开来,一直难以弄清。
多位研究人员的共同努力,促成了”睡眠中的神经优化(neural optimization in sleep)“这一概念的提出(隐喻性解释见下一节:Disk and RAM metaphor)。György Buzsáki 博士关于记忆痕迹形成的两阶段模型的开创性理论,为睡眠中究竟发生了什么投下了新的光芒(Buzsáki,19894)(重要提示:不要把这个两阶段模型与记忆的双组分模型(two-component model of memory)(Wozniak 等,19955),或下面将讲到的睡眠调节双组分模型(Borbély,19826)混淆)。凭借他对神经网络的知识、对神经元放电的精巧实验,以及对时空神经元放电模式的复杂数学分析,Buzsáki 提供了一个很好的模型,解释了睡眠的两个组成部分——REM 睡眠和 NREM 睡眠——如何协同工作以优化记忆。海马体充当大脑的中央交换机,能够轻易存储短期记忆模式。然而,这些模式必须被编码到新皮层(neocortex)中,以便为编码新的短期记忆腾出空间。这一重建大脑神经网络的复杂过程,就发生在睡眠期间。与休息或节能不同,这一演化神经数学的最高成就,要求大脑完全关闭与外界环境的输入(在大多数动物中)!这种自动的重新布线,正是我们睡眠的主要原因,也是没有意识处理参与其中的原因!在睡眠中,大脑工作得和参加 SAT 或 GRE 考试一样辛苦。它重新连接自己的回路,以确保所有新获得的知识都被最优地存储,以备将来使用。
我们睡眠,是为了让大脑能够整合新知识并形成新的联结。由于我们必须睡眠大脑才能继续运作,我们的身体也因此把许多重要过程挂接到睡眠中运行。简而言之,在清醒时我们使用和消耗,在睡眠时我们恢复和合成。睡眠影响着整个身体的功能与健康。
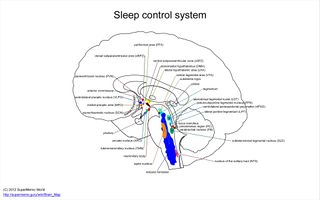
睡眠由一组复杂的脑核团控制。干扰这个系统会破坏睡眠的功能。详见:Sleep control system。
延伸阅读:
- 睡眠中的神经优化(Neural optimization in sleep)
- 并非所有科学家都同意(Not all scientists agree)
硬盘与内存的隐喻(Disk and RAM metaphor)
一个隐喻有助于理解睡眠的作用,以及为什么闹钟是有害的。我们可以把大脑及其 NREM-REM 睡眠周期比作一台普通的个人电脑。白天,在学习和体验新事物的过程中,你把新数据存储在内存(RAM)里。夜间,首先在 NREM 睡眠中,你把数据写入硬盘(hard disk)。接着在 REM 睡眠中(它在夜间紧随 NREM 之后),你进行磁盘碎片整理(disk defragmentation),即整理数据、排序、建立新的联结等。整夜之间,你反复进行”写入—碎片整理”的循环,直到所有内存数据都被整齐地写入硬盘(以供长期使用),而你的内存被清空,准备好迎接新一天的学习。醒来时,你重启电脑。如果你用闹钟过早重启,往往会留下磁盘碎片。你的数据访问变慢,思维混乱。更糟的是,有些数据可能根本没被写入硬盘,就像你从一开始就没把它存进内存一样。总而言之,如果你使用闹钟,你就在拿自己的数据冒险。如果你不在乎自己的智力表现,那你也许想知道,使用闹钟不健康还有许多其他的生物学原因。许多人使用闹钟并活了下来。但这和吸烟、滥用毒品、或沉迷于油腻的肥猪肉没什么两样。你可以用酒精虐待大脑多年,最终仍当上总统。人类的许多成就都需要被打断的睡眠。许多发明是由困倦的大脑创造的。但没有什么能像用一剂健康的、令人神清气爽的睡眠所刷新的大脑那样改变未来。
坏睡眠会致命,并耗资数十亿(Bad sleep kills and costs billions)
睡眠剥夺是杀手!它通过空难、核电站故障、车祸、石油泄漏等夺走宝贵的生命。睡眠剥夺可以改变历史的进程。如果 Charles Lindbergh 没能从一次因微睡眠(microsleep)引发的俯冲中恢复 Spirit of St. Louis(圣路易斯精神号),他将只是历史的一个脚注。睡眠剥夺改变了核裂变的未来,也改变了石油勘探的未来。糟糕的睡眠在路上夺走的人命和酒精一样多。在美国,每年有 1550 人死亡可归因于疲劳驾驶。这相当于十年里六起世贸中心倒塌的悲剧!令人惊讶的是,由于痛苦和苦难被稀释在整个人口之中,疲劳驾驶制造的新闻头条远不及一次恐怖袭击。至少三分之一的美国人曾在方向盘后睡着过!春季切换到夏令时(DST)时,车祸增加 9%。睡眠剥夺给工业化社会带来了天文数字的成本。学校里有数不清的小时被浪费在低效的学习上,有数不清的人工时被浪费在床上辗转反侧。暴躁的行为和尖锐的发脾气也有其成本。对更好睡眠的追求催生了绝望的解决方案,比如 Uberman 多相睡眠、“安全闹钟”装置、数百本书和数千篇关于快速入睡、早起、或少睡的博客。与此同时,真正的解决方案却简单而显而易见!阅读本文的相关部分,尝试至少一个月的自由睡眠,把你对睡眠及其改善你生活潜力的认知翻上四倍。我们需要敬畏睡眠,让孩子们睡够,设计更智能的夜班排班,并把那些关乎生死的职业(如医疗行业)中的睡眠剥夺降到最低。
在《经济学人》(The Economist)组织的一场睡眠剥夺辩论的结论评论中,Karen M. 写道:“我们睡得不够,而我们也不会’改变我们的方式’,因为在大多数人的一天里,做他们喜欢的事情的时间已经太少了。把这就称作生活中一个悲哀的事实吧,因为它本就是如此。” 尽管 Karen 试图用”我们”来代表整个人口,但本文的许多读者会不同意,并尽最大努力获得生理上必要的睡眠。否则我的写作努力就没有必要了。好睡眠让我们更友善、更聪明,还能拯救生命!
参见 WebMD 的 10 Things to Hate About Sleep Loss。
不睡觉,你就会死!(If you do not sleep, you die!)
几乎每个人都曾熬过通宵。即便这通常是一次不愉快的经历,它几乎总能在好好睡一晚后 100% 恢复。因此,知道一两周的睡眠剥夺就可能导致死亡,多少令人惊讶!睡眠研究人员制造了一种残酷的装置:一旦大鼠睡着就把它弄醒。这种装置表明,用睡眠剥夺杀死一只大鼠平均需要 3 周(仅剥夺 REM 睡眠则需约 5 个月)(Rechtschaffen,19987)。Siegel 博士证明了被剥夺睡眠的大鼠存在脑损伤(Siegel,20038)。由于糖皮质激素(glucocorticoids)水平升高,大脑某些区域的神经发生(neurogenesis)会被睡眠不足所抑制9。简而言之,睡眠剥夺对大脑健康非常有害。
睡眠剥夺是一种众所周知的酷刑形式。然而,出于伦理原因,大鼠实验无法在人类身上重复到其最终结局。不过,由于我们可以研究睡眠障碍的影响,我们对人类在睡眠剥夺状态下的耐受力有一个大致的概念。其中之一是致死性家族性失眠症(fatal familial insomnia):一种基因突变导致患者出现进行性加重的失眠,最终在几个月内死亡。另一个例子是 Morvan 综合征:一种自身免疫性疾病破坏神经元钾通道,导致严重失眠和死亡(除非病情进入缓解期)。
你可能听说过有人声称自己根本不睡觉的报告。这些肯定是不准确或虚假的。那些声称从不睡觉的人,要么是在吹嘘,要么是经历了一种睡眠状态知觉错误(sleep state misperception),使他们在床上休息时产生自己没有睡觉的错觉。
大脑的垃圾回收(Brain’s garbage collection)
为什么睡眠剥夺会致命?睡眠剥夺导致的死亡,就像整体上的衰老死亡一样。往往是多种原因共同作用,产生那个不可避免的最终结局。可能没有人确切知道这个谜团的答案。然而,对睡眠作用的研究给了我们相当强的提示。睡眠最重要的功能之一,是重组大脑中的神经网络。白天,我们学习新事物、记忆、习得技能、弄清问题、通过创造性联想建立新的记忆等。经过漫长一天的清醒,大脑中充斥着需要与先前所学整合的零散信息。没有这种重组,大脑会陷入混乱,并很快耗尽存储新记忆的空间。睡眠的这种神经作用如此根本,以至于睡眠剥夺几乎影响所有由神经系统支配的身体功能。没有定期的”垃圾回收”,各个网络开始出现故障。这些最初轻微的故障会累积成整个机体的严重问题。睡眠剥夺最显著的影响包括:体温调节问题、免疫功能下降、激素变化(如糖皮质激素和儿茶酚胺升高)、代谢变化[见:睡眠与葡萄糖代谢]、营养不良、幻觉、自主神经系统功能障碍、细胞粘附变化、炎症因子增加(如 IL-6、TNF、C 反应蛋白等)、皮肤损伤、氧化应激、DNA 损伤等。这些问题会严重到致命。打个比方,如果我们将一个较不发达的生物比作一架一战轰炸机,那么可以想象,演化成人类就像获得了驾驶一架 B-2 轰炸机所需的软件。尽管 B-2 比 Orville Wright 时代制造的飞机先进了不知多少代,但只要在其软件里植入一个 bug,就足以让它从天上掉下来。睡眠剥夺中的人体就像一架软件逐渐失灵的 B-2。它技术先进、它聪明,然而它也非常脆弱。依赖先进软件或神经功能总是危险的!幸运的是,消除这种危险我们只需要每天去睡觉。详见:睡眠中的神经优化。
睡眠保护(Sleep protection)
睡眠剥夺中还有第二层麻烦。由于睡眠的重要性,所有高等生物都实施了一套睡眠保护程序(sleep protection program)。这套程序确保睡眠剥夺会带来令人不快的症状,它还产生一种极其强大、难以克服的睡眠驱力(sleep drive)。保持清醒变得无法忍受,闭上眼睛成了宇宙中最令人感到安慰的事之一。这些症状是网络故障的结果吗?绝对不是。如果它们是,那么睡眠驱力本身也可能失灵。而且,从睡眠剥夺中的恢复也不会那么快、那么容易、那么彻底!睡眠保护程序确实存在,它能让睡眠剥夺的影响更糟。就像过度活跃的免疫系统中的细胞因子风暴一样,睡眠保护程序可能加剧由网络故障造成的损害。
合成代谢性睡眠(Anabolic sleep)
最后但同样重要的是,睡眠已演化成为机体主要的合成代谢(anabolic)状态。没有它,身体一直在消耗自身,几乎没有时间重建。开启合成代谢状态并不需要关闭意识,然而夜间休息的时间似乎是身体进行所有重建的最佳时机。由于我们反正必须睡眠,合成代谢功能与其他睡眠功能整合在了一起,现在可能已不可或缺。合成代谢状态,以及夜间 GH(生长激素)或睾酮的升高,也影响着神经网络和我们的”心智软件”状态。激素变化刺激和/或抑制神经生长。以证明睡眠在脑发育中作用而闻名的 Michael Stryker 博士(Stryker 等,200110)说,夜间激素变化可能”在巩固和增强清醒经验中扮演至关重要的角色”11。睡眠剥夺的一个主要死因似乎是由免疫功能下降(例如没有发热反应)导致的机会性细菌感染。这种下降同样可能由 (a) 免疫功能的神经控制不良,或 (b) 过度分解代谢(hypercatabolism)的直接效应造成。无论原因如何,科学家很快发现,使用抗生素对预防这些感染导致的死亡帮助不大。被剥夺睡眠的大鼠还是会死。感染可能只是加速了原本不可避免的死亡。
没有睡眠,我们为什么会死?
无法量化这三个因素对长期睡眠剥夺致命结局的贡献:
- 网络故障(network malfunction),或
- 睡眠保护程序的次级效应,或
- 持续的分解代谢(catabolic)状态。
即便后两者有可能通过药物手段得到缓解,也无法绕过睡眠中的网络重塑。那些希望找到对抗睡眠之解药的研究者,走的是一条死胡同。除非出现一些近乎科幻的严肃纳米技术,否则睡眠在人类身上还会存在很多年。尽管睡眠剥夺可能致命,睡眠仍是好消息。它让我们更聪明!我们都应当拥抱健康、不受约束的睡眠所带来的福祉。毕竟,生活中很少有比在充实的一天后睡个好觉更好的事了。睡眠应被列入基本人权!
睡眠的两个组成部分(Two components of sleep)
电灯和压力是把自然的睡眠过程变成数百万人日常挣扎的两个主凶。在新的千年里,如果不理解睡眠的科学和艺术,我们很少能指望获得好眠。目前社会对睡眠及其功能的理解,就像 20 世纪 20 年代对香烟健康风险的理解一样糟糕。大多数人通过用闹钟、不合理的轮班模式、安眠药、酒精、咖啡因等来调节睡眠,从而给自己和孩子带来痛苦、悲惨和精神折磨。
要打破不健康的睡眠习惯,你需要理解睡眠调节的双组分模型(two-component model of sleep regulation)。
驱使你上床的嗜睡感有两个组成部分:
- 昼夜节律组分(circadian component)——嗜睡感以大约一天长度的周期循环回到我们身上。
- 内稳态组分(homeostatic component)——嗜睡感随我们保持清醒的时间长度而增加。
只有这两者的结合,才能确定最佳的睡眠时间。最重要的是,你应该记住:即使由内稳态组分产生的强烈嗜睡感,如果时机与昼夜节律组分所决定的最大睡眠倾向相悖,也可能不足以获得好睡眠。
昼夜节律组分(Circadian component)
大约有一百种已知的身体机能,在一天长度的周期里于最大值和最小值之间振荡。由于这些机能完成一个周期大约需要一天时间,1959 年德国科学家 Franz Halberg 博士创造了昼夜节律(circadian rhythm)一词(拉丁语 circadian 意为”大约一天”)。维持睡眠的总体倾向也受这种昼夜节律支配。在一般情形下,最大嗜睡感出现在半夜,在醒来时降至最低,并在下午午休(siesta)时间再次小幅上升。然而,昼夜节律的嗜睡感常常与你期望的睡眠时间在相位上存在偏差。因此,如果你的最大嗜睡感出现在早晨,你可能会发现即使在深夜也很难入睡,哪怕前一天你错过了大量睡眠。换言之,你的最佳睡眠时机应当考虑到你的昼夜节律。
内稳态组分(Homeostatic component)
内稳态(homeostasis)是指维持生理和代谢功能平衡或均衡的术语。如果你喝下含有大量钙的液体,内稳态机制会确保你通过尿液排出钙或将其沉积在骨骼中,以确保血液中的钙水平保持不变。类似的机制也被用来调节整体的嗜睡感及其多个子成分。你清醒的时间越长、学习越多、思考越多,你入睡的倾向就越高。另一方面,咖啡因、压力、运动等因素可能暂时降低你的内稳态嗜睡感。内稳态机制在一天繁重的脑力劳动后为你入睡做好准备,同时在紧急情况下阻止你入睡。
时钟与沙漏的隐喻(Clock and Hourglass metaphor)
一个隐喻有助于解释睡眠的两个组成部分(更科学的解释见:Borbély model)。在大脑深处,你的生物钟运行着一个 24 小时的活动周期。每 24 小时,生物钟会隐喻性地释放一剂”催眠药水(sleepy potion)“让你入睡(详见:Why we fall asleep)。如果你试图在错误的时刻、没有这剂催眠药水的情况下入睡,可能会发现非常难睡着。所有失眠症患者都苦于缺乏催眠药水。如果他们在获得那剂药水之前就过早入睡,他们会在床上辗转反侧,常常长达数小时。你需要倾听自己的生物钟,才知道何时该去睡觉。
重要的是要知道,生物钟产生的催眠药水不足以让你入睡。大脑还使用一个”精神能量的沙漏”,它每天给你一定的时间用于脑力工作。你醒来时沙漏是满的,并开始被倒空。每一个清醒的瞬间、大脑吸收的每一点信息、每一次脑力付出,沙漏都越来越不满。只有当精神能量的沙漏被倒空,你才能快速入睡。
要获得一夜好眠,你需要结合两个因素:
- 你的生物钟必须说”该睡觉了”(睡眠的昼夜节律组分)。
- 你的能量沙漏必须说”再也干不动脑力活了”(睡眠的内稳态组分)。
如果你的催眠药水试图让你入睡,但你的精神能量沙漏还是满的,你会非常昏沉、疲倦,却睡不着。另一方面,如果你在没有催眠药水的情况下试图入睡,而能量沙漏已空,你可能成功,但你会很快醒来,而此时沙漏又满了,这让再次入睡几乎不可能。失眠症患者是在生物钟释放催眠药水之前就去睡觉。当你被闹钟早早叫醒时,你几乎站不稳,因为体内充满催眠药水,它恳求你回去继续睡。当你在下午昏昏欲睡时,你的精神能量沙漏可能几乎空了。一次快速的小睡能帮你把它重新填满,从而在晚上非常高效。如果你在早上喝咖啡,它帮你给沙漏充能并增加一些额外的精神能量。但咖啡与催眠药水结合,会产生一种有毒的混合物,让你的大脑笼罩在病态的瘴气中。如果你试图在夜里靠喝咖啡保持清醒,你会感觉像被马踢中了胃部。那是对大脑健康最彻底的犯罪式攻击。
好睡眠的基本定理(The fundamental theorem of good sleep)
现在让我们来表述好睡眠的基本定理:
要获得能最大化学习效果的高质量夜间睡眠,你的入睡时间应满足两个条件:
- 强烈的内稳态嗜睡感:这通常意味着,距离从上一夜睡眠中醒来不少于 15-19 小时才入睡。
- 上升中的昼夜节律嗜睡感:这意味着在你通常经历困意迅速上升的时刻入睡。不早不晚!了解你昼夜节律的时机对一夜好眠至关重要。
你应该意识到,利用昼夜节律组分只有在它所有的生理子成分同步运行时(即在自由睡眠状态下)才能奏效。睡眠时间不规律、生活高度紧张的人,可能根本无法定位”上升中的昼夜节律嗜睡感”这一点,因为这一点可能根本不存在!关于昼夜节律和内稳态组分的可视化说明,见”SuperMemo 中的双组分睡眠模型”一节。关于睡眠两个组成部分的更多内容,见:Borbély model。
好睡眠什么时候可能不来?
你可能会惊讶地发现,你内在的昼夜节律振荡,所基于的周期更接近 25 小时而非 24 小时!准确地说,它在不同个体、季节以及压力、睡眠时机、光照时段、光照强度、运动等日常因素之间变化,通常落在 24.5 到 25.5 小时的范围内。
我们大多数人都能通过使用重置振荡的因素,把这个约 25 小时的昼夜节律驯化为 24 小时周期。这些因素包括强烈的晨光、工作、运动等。德国科学家把这些因素命名为授时因子(zeitgebers,即”给予时间”的因素)。在授时因子的影响下,一个调整良好的个体,其周期每天可以被拨回 30-60 分钟。然而,对许多个体来说,驯化到 24 小时周期可能很困难,原因包括:
- 失明(即无法使用主要的授时因子:光)
- 近视(即对光授时因子敏感度降低)
- 睡眠需求增加(例如高强度学习、高度创造性的工作、运动等的结果)
- 压力
- 内分泌失调
- 睡眠障碍
- 青春期
很大一部分睡眠障碍可以用驯化失败(entrainment failure)(即未能把 25 小时的昼夜节律重置到 24 小时的日周期)来解释。换言之,在睡眠障碍与驯化失败的相互依赖中,因果关系常常是颠倒的!由于睡眠的生理功能是重新布线大脑的神经网络,我们自然可以预期,睡眠需求与前几天学习量相关联。这种联系也可能解释退休后由于脑力活动减少而导致的睡眠需求下降。在高度活跃的个体中,这种与年龄相关的睡眠需求下降较不可能被观察到。出于类似原因,驯化失败在考试期间的学生中常常可以被发现。其中有多少归因于压力、有多少归因于想在一天里做更多事、又有多少归因于睡眠需求的实际增加,尚不清楚。
好睡眠的公式(Formula for good sleep)
有一个鲜为人知的公式,能完美治愈那些持续或季节性受睡眠驯化(entrainment)问题困扰的人。这个公式就是自由睡眠(free running sleep)!
自由睡眠的定义是:戒除一切形式的睡眠控制,如闹钟、安眠药、酒精、咖啡因等。自由睡眠是在内稳态组分和昼夜节律组分共同触发下,在内部自然触发的时刻来临的睡眠。换言之,自由睡眠意味着:只有当你真正困倦时(无论此刻与一天中的什么时间有何关系)才去睡觉。自由睡眠作息下的夜间睡眠,持续时间由身体需要决定,并以自然醒来结束。任何形式的睡眠中断都不被允许。尤其是,使用闹钟是对自由睡眠原则的根本违背。
自由睡眠最大的缺点是,它常常会导致周期长于 24 小时,这使其难以在社会中广泛使用。然而,如果你想尝试自由睡眠,或许可以在假期进行。在理解你的昼夜节律之前,你可能需要一个持续两周以上的假期。即便你在非假期环境中负担不起自由睡眠,尝试一次也会极大增进你对自然睡眠周期,尤其是你自己周期的了解。你也应知道,可以把睡眠驯化到一个期望的时段(例如早起)。然而,驯化需要铁一般的自律,以及对驯化规则一丝不苟的遵守。
自由睡眠(Free running sleep)
自由睡眠是指不被人为控制以迎合我们日程和愿望的睡眠。它是一种没有闹钟、没有安眠药的睡眠。人类自存在以来就在实践自由睡眠。我们的祖先在日落时被温柔地劝诱上床,并自然醒来,很可能在床上度过不少于 8-10 小时(另见:Segmented sleep)。所有偏离这一健康做法的做法,都是文化、习惯、宗教和/或传统的强加。尽管我们祖先的生活充满危险、迷信、战争和疾病,我们仍应停下来想想,这种自然不被打扰的睡眠对他们健康的奇妙影响。火和烛光的出现并未提供太多熬夜的动力,除了少数晚上总有许多事可做的人:最早的”书虫”和艺术家。只有爱迪生等人的天才带来了真正的睡眠灾祸:电。随着印刷品和电照明的广泛传播,数百万人发现晚上的书比睡眠有趣得多。然后是网络。到了 2012 年,我们有一整谱系无穷无尽的娱乐和消遣,诱惑每个人离开床和健康的睡眠。我们越来越想把睡眠挤进”设计师定制的时段”里。我们希望在特定的时间入睡,在特定的时间醒来。令人惊讶的是,很大一部分人没有意识到:不损害健康,这是不可能的!幸运的是,几乎每个人都有”睡眠对健康生活至关重要”的直觉。那些想彻底不睡的人只占极少数。几乎所有创造型人士都希望醒来时神清气爽、随时可以行动,最好是在指定的时间。同样的人也希望晚上睡前没那么累,并能瞬间入睡,最好也是在指定的时间。那么让我用粗体郑重声明:
如果排除不健康的技巧:
- 我们不可能随心所欲地入睡。
- 我们不可能随心所欲地醒来。
- 我们不可能消除晚上的困意。
尽管这令人失望,但每个人若能消化这些真相,都会在生活中做得更好。如果我们同意在身体偏好的时间自然醒来,那么从醒来的那一刻起就保持神清气爽应是可能的。然而,清醒的一天中脑力能力的下降是不可避免的,这是自然的。中午警觉性的低谷也是不可避免的。而最佳的入睡时间几乎不可移动。如果你试图提前它,你很可能会经历失眠。如果你试图推迟它,你会减少睡眠并可能醒来时没有精神。如果你试图比自然时间更早醒来(例如借助闹钟),你会在一定程度上以睡眠剥夺的状态醒来,这会削弱睡眠对你学习和创造力的价值。不要被闹钟带来的警觉性幻觉所迷惑。是的,这对某些人、某些时候会奏效,这助长了”靠闹钟铃声能更神清气爽地醒来”的迷思。
健康、令人神清气爽的睡眠只有一个公式:只有当你非常疲倦时才去睡觉。不早不晚。没有闹钟自然醒来。
这个简单的公式叫做自由睡眠。对许多人来说,在多年的睡眠滥用之后,即便是自由睡眠也可能很棘手。发现自己身体的规律并接受它们,需要一段时间。如果你能正确执行自由睡眠,你会知道:入睡不超过 5 分钟(不借助药物、酒精或其他干预),并以神清气爽的感觉相当突然地醒来。早晨神清气爽不能想当然。即便睡眠与昼夜节律相位有轻微的错位,也会夺走这种神清气爽的感觉。在数周或数月混乱的睡眠之后,一些昼夜节律变量可能在不同周期中运行,自由睡眠不会是立竿见影的解药。把它调节到足够好以达成目标可能需要一些时间。甚至不能排除,经过多年的轮班工作或时差,睡眠控制中心的一些脑细胞可能已经死亡,使得获得良好对齐、令人神清气爽的睡眠更加困难。除这些注意事项外,压力是破坏睡眠肌理的主要因素之一。在自由睡眠中,压力会让你睡得更晚、入睡更难、醒得更快且远不那么神清气爽。为了长寿和生产力,对抗压力是每个人生活中最重要的事情之一。
伴侣和配偶可以各自在独立的周期中自由睡眠,但他们常常会惊讶地发现,彼此之间比与世界其他部分更容易同步(尤其是当他们有相似的兴趣和日常作息时)。如果他们同床共眠,通常其中一人会稍早起床,并对另一人充当强有力的授时因子。只有当两人自然偏好的睡眠周期长度差异显著时,问题才会出现。在这种情况下,对方不再是授时因子,而是闹钟的替代品。
即便你不信服,你也应当尝试自由睡眠,以更好地理解睡眠相位(sleep phase)的概念,以及各种生活方式因素如何影响睡眠相位。你常常会注意到,你所谓的睡眠障碍消失了!请注意,自由睡眠的周期并不完全是遗传的。日常作息中的各种因素能缩短或延长周期。显然的包括:晨光或在傍晚服用褪黑素可缩短周期;傍晚令人兴奋的活动会延长周期。周期随季节略有变化,度假时也会改变,并常随年龄增长而变短。尝试自由睡眠以了解你自己的睡眠参数,这将帮助你与世界同步,或至少获得令人神清气爽的高质量睡眠。请在本文中阅读更多关于自由睡眠的内容。扔掉闹钟不是万灵药,你可能需要学一点睡眠卫生(sleep hygiene)。
我们应该自由地睡眠吗?(Should we free run our sleep?)
如后文所述,自由睡眠可用于解决许多睡眠障碍,除了那些破坏自然睡眠机制的潜在器质性障碍。然而,你常会听到两个反对采用自由睡眠的论点:
- 论点 1——自由睡眠常常会导致一天长于 24 小时。这最终会导致在不寻常的时间睡觉。这似乎违背了自然的 24 小时光暗周期。较少见的情况下,周期会短于 24 小时。
- 论点 2——睡眠可以比作进食。你的身体总会试图获取比实际需要更多的睡眠。这会导致在睡眠上花费不必要的时间。换言之,自由睡眠是低效的。
论点 1:相位移动(Phase shifts)
自由睡眠确实常常会与自然的光暗周期相悖。然而,偏离自然节律是使用电照明和现代生活方式的直接后果。我们的祖先在日落之后除了黑暗和无聊之外,几乎别无所求。漫长的黑暗和无聊在哄人入睡方面相当有效。如果我们固执地拒绝在某个时刻之后使用电灯,我们仍会发现很难摆脱现代生活方式的种种刺激。要在大脑能在傍晚早早入睡,你大概需要辞去现在的工作,找一份乏味的工作,放弃紧张的家庭生活,放弃爱好和兴趣,放弃互联网、晚间电视等。我们比祖父母过着压力更大、更刺激的生活。在傍晚早早点灯可能只是浪费。此外,作为信息时代通病的近视,使我们对光授时因子更不敏感,人为地延长了昼夜节律周期。自由睡眠有一些缺点,最糟糕的是难以建立一个与世界其他部分良好同步的活动周期。通过自律遵守周期重置规则(如晨练、强光、晚间睡眠保护区等),周期的稳定是可能的。
论点 2:过度睡眠(Excessive sleeping)
尝试自由睡眠的人,在最初确实可能发现自己睡得离谱地长。然而,这对健康个体来说不会持续,因为长睡眠是身体对睡眠剥夺所致各种睡眠亏欠的代偿反应。与食物不同,在我们能够睡得更长的日子里获得额外睡眠,似乎没有任何演化上的优势。在演化过程中,我们发展出了过度进食的倾向,这是对食物匮乏时期的保护。脂肪组织充当了艰难时期的生存储备。然而,考虑到睡眠的功能,睡眠需求应与前几天的学习量大体成正比。在古代,我们没有考试日与清闲日的对立。因此,学习与睡眠需求之间的联系相当弱。即便在整天无所事事之后的日子,生物钟仍会让我们在夜里睡 7-8 小时。其次,每多睡一分钟都可能改善大脑神经布线的质量。睡眠最好比作饮水而非进食。由于我们糟糕的水分储存能力,我们没有多少不喝水也能存活的能力。同样,我们无法提前睡眠,为考前或重要截止日期前的双重通宵做准备。“自由睡眠会增加对睡眠的自然需求”这一说法是错误的!如果你在自由睡眠中睡得更长,这说明你在自由睡眠之前处于睡眠剥夺状态。这个长睡眠阶段是短暂的。 偶尔,如果你睡得过早,也会累积异常多的睡眠时长。详见:Excessive sleeping。
在我看来,每个人都应当始终自由地睡眠,除非这使其无法按照所选职业、专业、教育等在社会中运作,或使其无法照顾年幼的孩子。
自由睡眠是无压力的(Free running sleep is stressless)
有人提出,如果任何改变都带来压力,那么转向自由睡眠也会有压力。情况恰恰相反。或许除了最初的调整期(在这个阶段,对时间生物学了解较少的人会犯错,可能导致睡眠质量下降)。说任何改变都有压力,是一个走得太远的概括。每天换 T 恤并不意味着压力。此外,改变的程度很重要。同样的改变,根据其程度,可能造成过度压力,也可能是生活中受欢迎的因素。让你的睡眠自由运行并不意味着任何程度的压力,除非自由睡眠本身在你日程中产生了可能带来压力的变化。如果你在感到饥饿时适度频繁地进餐,你可能比在预设的午餐时间进餐体验到更少的压力变化。根据定义,自由运行的行为释放你的机体,使其将行为适应身体的内部需求。因此,这些可被视为抗压力因素。这同样适用于睡眠、饮食习惯、运动和其他生理需求。
自由睡眠算法(Free running sleep algorithm)
-
从一份一丝不苟的日志开始,记录你入睡和早晨醒来的时间。如果白天有小睡,也记入日志(哪怕小睡只有 1-3 分钟)。日志将帮助你预测最佳睡眠时间并改善睡眠质量。一旦自我研究阶段结束,你将积累足够的经验,不再需要日志;但在开始时你需要它来更好地理解自己的节律。你可以使用 SleepChart 来简化记录流程,并帮助读取你的昼夜节律偏好。
-
只有当你真正疲倦时才去睡觉。你应能感觉到你的睡眠潜伏期(sleep latency)可能少于 5-10 分钟。如果你没有把握能在 10-20 分钟内入睡,就不要去睡!如果这需要你熬夜到凌晨,那就熬吧!
-
确保没有任何东西打扰你的睡眠!不要使用闹钟!如果可能,在没有伴侣的情况下睡眠(至少在自我研究期)。让自己与噪音源和光照的快速变化良好隔绝。
-
白天,尤其是傍晚,要避免压力。这在自我研究期尤为重要,此时你仍不确定自己的最佳睡眠模式是什么样。压力激素对睡眠的时机有强大影响。在你本该入睡的时刻,压力性的想法也可能让你保持清醒。
-
几天后,尝试算出你昼夜节律周期的长度。如果你得到的数字大于 24 小时,你的自由睡眠会导致每天越来越晚入睡。这最终会让你在白天睡觉。这就是为什么你可能需要一个假期来认真测试自由睡眠。一天长于 24 小时是相当正常的,你可以通过恰当定时的信号(如光照和运动)来稳定模式。如果你是 DSPS 类型,这可能非常困难。
-
一旦你知道自己平均保持清醒多长时间,就每天计算你预计入睡的时间(我用**预计入睡时间(expected bedtime)和预计就寝时刻(expected retirement hour)**这两个术语来表示上床时间和入睡时间,在自由睡眠中二者几乎相同)。这一计算将帮助你预测入睡时刻。某些天你可能在预计入睡时间之前就感到困倦。不要与困意对抗,去睡觉,即便这比你预计的入睡时间早 2-3 小时。同样,如果在预计入睡时间你不感到困倦,就保持清醒、保持忙碌,晚些再睡,即便这比你预计的入睡时间晚 2-4 小时。
自由睡眠中的致命错误(Cardinal mistakes in free running sleep)
-
不要在足够困倦之前就去睡觉——这可能导致入睡 10-30 分钟后醒来,然后醒着 2-4 小时。最终你可能经历整个周期的人为前移!
-
除非有自然原因(不困),否则不要在远晚于预计入睡时间之后才去睡觉。这将导致错过最大昼夜节律困倦期。你的睡眠会更短、更不令人神清气爽。你的测量会更不规则,你会发现更难预测接下来几天的最佳睡眠时机。
-
不要在醒来超过 7-8 小时之后才小睡。晚的小睡可能会影响预计入睡时间并扰乱你的周期。如果你在傍晚感到困倦,你将不得不等待那个你认为能一觉睡到天亮的时刻。
睡眠记录提示(Sleep logging tips)
在自由睡眠条件下,记录实际睡眠时间不应困难。在驯化失败的条件下,你可能难以入睡,或”分阶段”慢慢醒来。在自由睡眠中,你应该能很快达到这样的状态:不到 10 分钟入睡,并立刻醒来(即没有睡眠惰性期)。换言之,你能记住上床的时间,加上 5-10 分钟,把它记为入睡时间。早晨一睁眼,你就应记录醒来时间。通常你不会有”是否已经彻底醒来”(相对于暂时醒来)的疑问,而且通常不会再睡着(在非自由睡眠中这可能是常见情况)。下图以图形形式展示了一份示例性的自由睡眠日志:
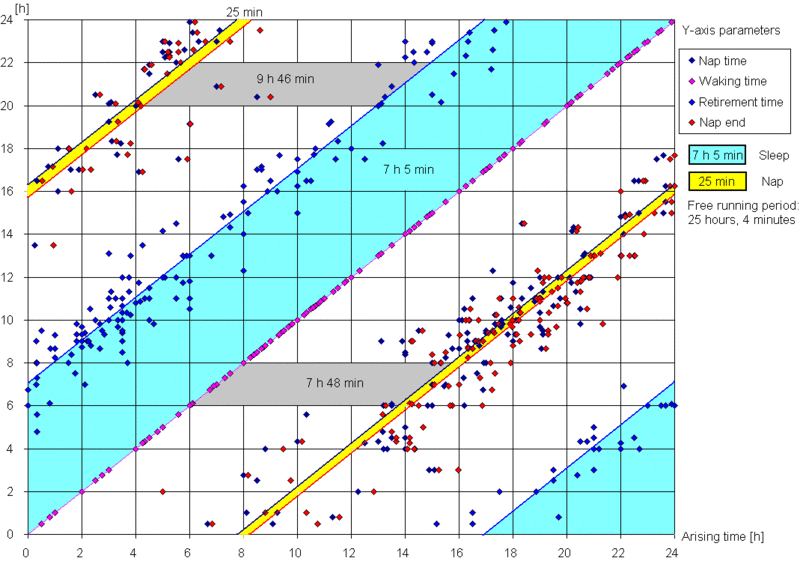
一份示例性的五个月自由睡眠周期图。图中夜间睡眠平均时长 7 小时 5 分,午睡前清醒时间 7 小时 48 分,平均午睡时长 25 分钟,午睡与夜间睡眠之间的时间 9 小时 46 分。整个周期合计 25 小时 4 分钟。注意图中午睡与夜间睡眠之间的距离小于 9 小时 46 分,这是因为蓝色的就寝线是相对于前一天睡眠而言的,而红色的午睡线则不是。因此,午睡-睡眠带在水平方向上缩短了 64 分钟,正好等于周期的每日相位移动量。
如果你用 SleepChart 收集了自己的自由睡眠数据,我将非常感谢你的提交,这些数据将对进一步研究有用(从 SleepChart 发送数据只需一次点击)。
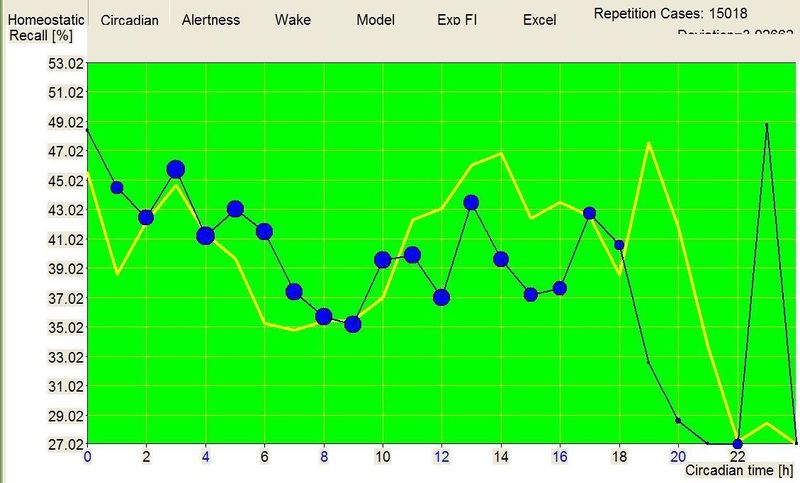
优化脑力工作的时机(Optimizing the timing of brainwork)
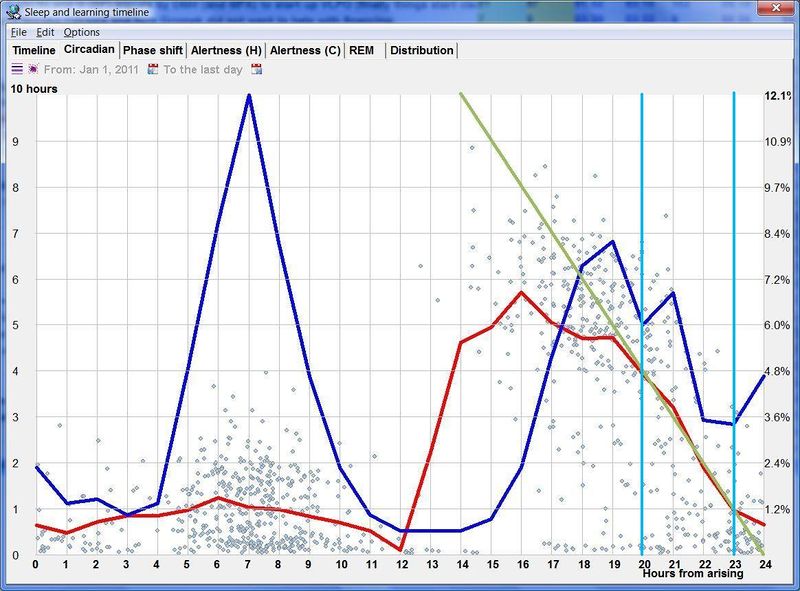
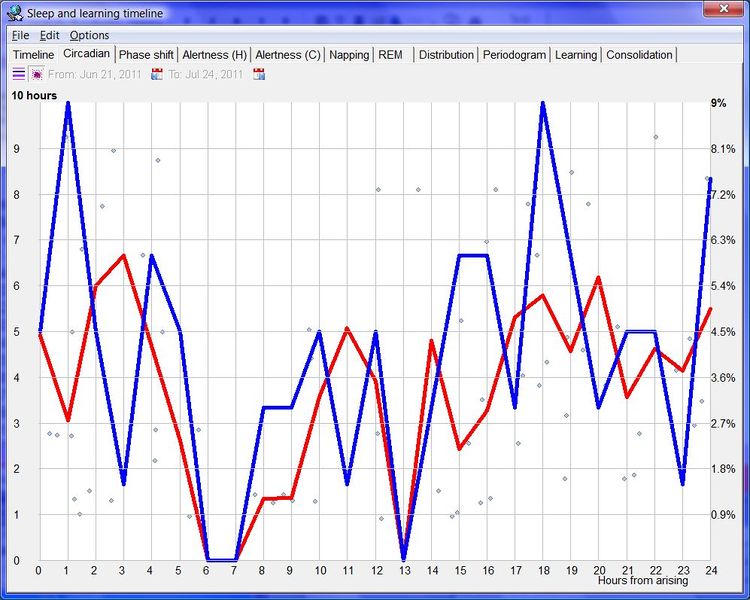
昼夜节律图与脑力工作
下面这张示例性的昼夜节律图,是用 SleepChart 基于一份自由睡眠日志生成的:
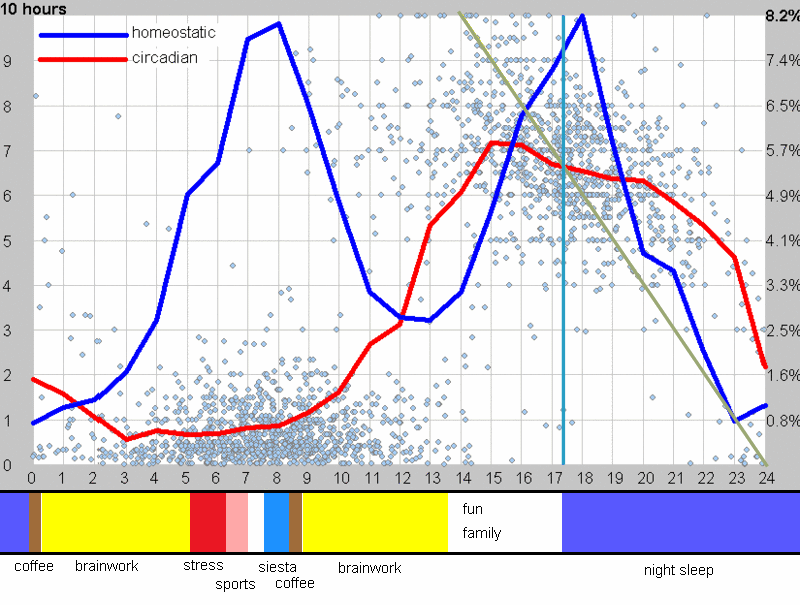
横轴表示从醒来开始的小时数(注意自由运行的节律周期通常长于 24 小时)。浅蓝色的点是实际的睡眠事件测量,横轴为时机,左侧纵轴为长度。内稳态困倦大致可用发起睡眠的能力来表达。已发起睡眠事件的百分比绘制为一条粗蓝线(右侧纵轴刻度)。内稳态睡眠倾向与脑力付出成正比增加,并可被咖啡因、压力等部分清除。昼夜节律困倦大致可用维持睡眠的能力来表达。已发起睡眠事件的平均长度绘制为一条粗红线(左侧纵轴刻度)。中午的警觉性低谷也是昼夜节律性的,但生物学上有所不同,导致的是不会在红色睡眠维持峰上登记出来的短暂睡眠。睡眠维持的昼夜节律组分与以下因素相关(但不相等):(1) 负相关于:体温、ACTH、皮质醇、儿茶酚胺,以及 (2) 正相关于:褪黑素和 REM 睡眠倾向。更多细节见:Circadian graph 与 Biphasic nature of human sleep。
最佳脑力工作时间(Best brainwork time)
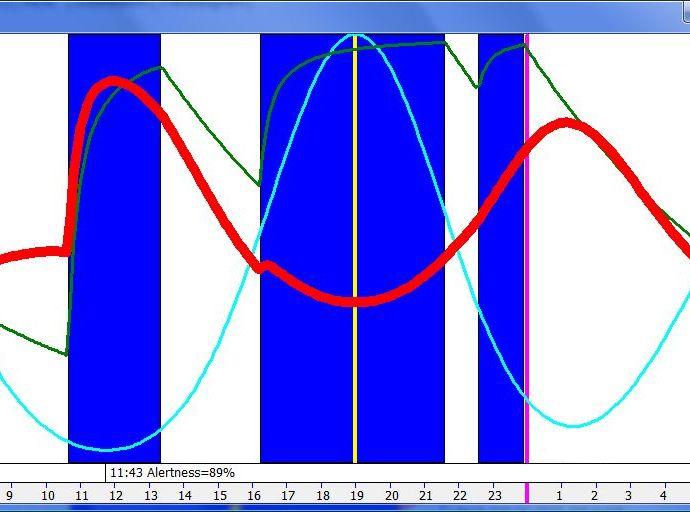
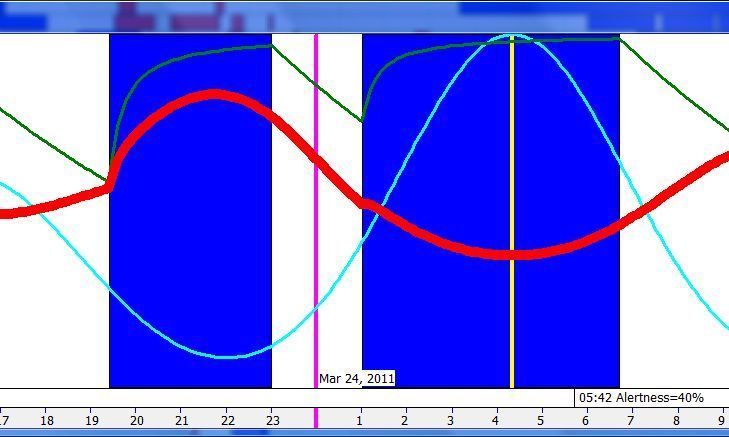
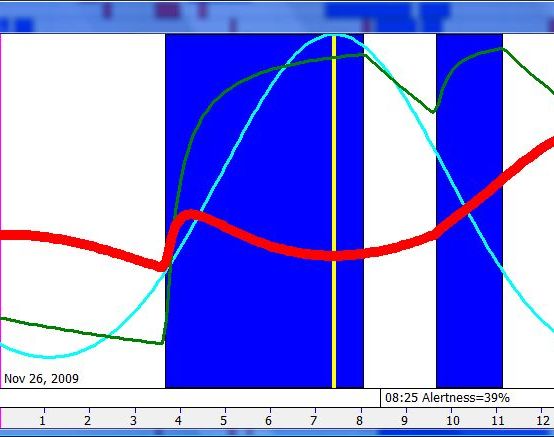
脑力工作的最佳时机,需要内稳态困倦和昼夜节律困倦都很低。一天中有两段高质量警觉时段:第一段在醒来之后,第二段在午休(siesta)时段之后。两段在图中都以黄色区块标出。为获得最佳学习和最佳创造性成果,请把脑力工作安排在这些黄色区块。咖啡因只能用于在这个最佳时段的早期增强警觉性,之后的使用将影响睡眠(咖啡因半衰期约 6 小时)。运动的最佳时机因你的运动目标而异,也取决于授时因子的最佳时机(例如,DSPS 型人适合清晨,ASPS 型人适合傍晚)。在本例中,压力区块之后跟着运动区块,以在午休前抵消压力的激素和神经效应。未标记的白色区域可用于午餐(午休前)以及晚上无关工作的娱乐时间,此时上升的昼夜节律困倦让创造性工作无效。那个白色的晚间保护区应远离压力、酒精、咖啡因等。推荐的活动可包括娱乐、游戏、放松、看电视、阅读、家庭、DIY、家务等。对那些积习已深的工作狂,挑战性较小、无压力的工作可能也行。**判断一天安排是否合理的最佳石蕊试纸是:所有活动都应是令人愉悦的!**脑力工作只有在你的大脑准备好时才令人愉悦。睡眠只有在准备好时才令人愉悦。休息和娱乐只有在充实的一天之后才恰如其分。甚至一点点压力,如果剂量和时机恰当,也可以是令人愉悦的。你不必做个肾上腺素瘾君子才能享受你的压力和运动时段。毫不夸张地说,对昼夜节律周期的良好理解,是拥有快乐而高效的一天的关键!
平衡的 24 小时周期(Balanced 24 hour cycle)
那条倾斜的绿线把图分成相位前移(右侧)和相位延迟(左侧)区域。这条线由图中那些”醒来时间(横轴)加上睡眠时间(左侧纵轴)等于 24.0 小时”的点确定。绿色盈亏平衡线与红色睡眠长度线相交的位置,决定了最佳的、平衡的 24 小时睡眠周期。在所给的例子中,17.35 小时的清醒加上预计 6.65 小时的睡眠,构成一个完整的 24 小时睡眠-清醒周期。绿线与红线之间的夹角越大,越难平衡睡眠并将其塞进地球自转的 24 小时周期。在本例中,增加清醒时间并不能让睡眠缩短到足以轻松实现平衡的程度。这意味着,可能需要一丝不苟地遵守 17.35 小时的清醒日来平衡周期。然而,这个缩短的清醒日可能增加睡眠潜伏期并增加过早醒来的概率,这也可能使平衡向相位延迟倾斜。垂直的青色线表示预计睡眠时间加上醒来时间等于 24 小时的位置(与代表完美 24 小时日的绿线相交)。在 DSPS 或 ASPS 中,这种 24 小时平衡可能难以达成。例如,在没有医疗干预的情况下,DSPS 要平衡一天,只能靠晚上一个大的保护区、早的小睡(或不小睡)以及高强度的晨练。
重要! 此图基于仅在自由睡眠条件下才成立的数据。如果你用闹钟来调节睡眠时机,这些测量和建议可能不适用!此外,时机的幅度变化因人而异,差异巨大!
违背自然节律地睡眠(Sleeping against your natural rhythm)
如果你违背自然节律睡眠,你常常会经历可通过调整睡眠时间来解决的疲倦或困倦。在健康个体中,白天的警觉性主要由以下因素决定:
- 昼夜节律相位和内稳态困倦
- 前一晚的总睡眠时间
- 前一晚的慢波睡眠(slow-wave sleep)量
- 前几天对睡眠-清醒作息的规律遵守
- 前几天累积的睡眠亏欠(例如 REM 亏欠、SWA 亏欠等)
所有这些因素都与睡眠相位密切相关。自由睡眠提供了在整个清醒一天中最大化警觉性的最佳方式。自由睡眠可能把最低体温点从清晨移向主观夜晚的中段。你应该注意到,在入睡前的困倦感增加,而在醒来时没有睡眠惰性!如果你无法自由睡眠,理解你的内稳态和昼夜节律睡眠驱力之间的关系(如下表所汇编)就非常重要。在一天之中,你应在表格的黄色区域之间同步移动,即从完美警觉到最大困倦,再回到完美警觉。灰色区域说明你的睡眠何时失步。
杀死闹钟!(Kill the alarm clock!)
闹钟流行病(Alarm clock epidemic)
在现代激烈竞争的社会里,很少有向上爬的人能不靠闹钟生活。靠一杯浓咖啡和全天候的压力,大多数人学会了带着闹钟生活和生存。一半的人口被闹钟叫醒,9% 被伴侣叫醒,4% 被宠物叫醒,3% 被孩子叫醒等,剩下自然醒来的只是少数。在一个以速度和完美时间管理为成就标志的社会里,时间越来越成为最珍贵的商品。然而,闹钟带来有害的副作用:压力、睡眠债,以及最糟糕的——破坏睡眠的自然生理功能。在最坏的情况下,这些因素会导致大脑的物理损伤(例如像海马体这样敏感的结构——你的记忆交换机——可能因睡眠被打断而实实在在地失去神经元)。
时间管理的艺术使人有可能带着身边的闹钟高速生活,同时免于压力。然而,闹钟和睡眠剥夺给社会造成的损害是不可原谅的。打断你睡眠的闹钟,损害你的记忆、你的学习能力、你的创造力、你的情绪和脾气、你与他人的关系、你的专注力,以及你整体的智力表现!
Robert Stickgold 博士已经证明,白天学习一项技能的人,直到获得 7-8 小时结构良好的充分睡眠后才表现出显著进步3。进步程度与所获睡眠质量之间存在明显相关性。我自己用 SleepChart 的工作也表明,使用闹钟会极大地降低记忆的回忆(recall)和巩固(consolidation)。遗忘是无痛的,以至于我们很少注意到它的影响。自然地,即便你睡得够多,遗忘也会发生,而要指出哪些具体记忆因睡眠不足而丢失是困难的。此外,睡眠剥夺可能让你的记忆完好,但其存储是次优的。这种差异在没有测量的情况下可能无法察觉。我们更可能注意到的是困倦、脑力迟钝或坏心情。
对睡眠的不尊重已达到圣经般的程度。这在美国及其他高度工业化的国家最为明显。《Men’s Health》的 Dan Vergano 为 ABC News 撰写的《No More Rude Awakenings》提出了一套对抗困倦的七天系统:“秘诀在于为那个唤醒系统提供燃料,让它能把睡眠系统打得落花流水。通过创造那种触发激素以唤醒大脑的良好预期,你将克服睡眠的需求,并能像着火的人一样跳下床。” 文章提出了一种利用”压力激素有助于保持警觉”这一事实的”清新头脑”法。然而,对于”粗暴唤醒”有一个简单且唯一理性的解药:睡够!像着火的人一样跳起来,不太可能对你的创造潜力产生积极影响!
你可能常常注意到,被闹钟叫醒能让你迅速开始一天。你可能因此相信,使用闹钟有助于你在白天晚些时候保持警觉。事实并非如此。闹钟信号只是把你的大脑吓得清醒过来,破坏了睡眠中精心规划的神经优化过程。结果,你立即获得一剂肾上腺素的注射,ACTH和皮质醇水平也升高。正是这种在自然睡眠节律中于醒来时达到峰值的皮质醇,给了你”清新头脑”的印象。随着时间推移,这种廉价获得的警觉性会消退,除非你继续用更多”疗法”虐待你的生理。你可能用更多恐吓手段保持警觉,滥用咖啡因,甚至用莫达非尼(modafinil)、可卡因或安非他命获得更强的效果。警觉性应通过充足的睡眠来获得,而不是在缺乏睡眠的情况下勉强为之!除了降低你学习新事物的能力外,所有非自然的抗困倦方法都会产生大量副作用,长期来看对你的健康可能相当有害。
所有试图用睡眠以外的方式来克服困倦的努力,都可以比作在使用精神活性物质时追逐第一次的快感。如果你喝大量咖啡、做俯卧撑、往头上浇冷水、或拍打自己的脸,你只是动用了你最后的警觉激素储备,而这只会让刺激效果消退后的剥夺效应更糟,这通常只是几分钟的事。很少能获得持续超过一小时的提升,而你越兴奋,随后的低谷就越深。
失眠陷阱(Insomnia trap)
如果你的生活没有闹钟似乎是不可能的,你可能需要用尽书中所有方法来确保获得足够的睡眠并把伤害降到最低。如果你需要以牺牲大脑为代价早起,请避免失眠陷阱(insomnia trap)!失眠陷阱是一个恶性循环:
- 为了多睡而过早上床,
- 未能及时入睡(或更糟,过早醒来),
- 第二天感觉更累,以及
- 第二天为了补回失去的睡眠而更早上床。
最好是按自然时间(稍晚一点)入睡,早起,承受一定程度的睡眠剥夺,并寄望于一次相位重置,让你能继续按设计师定制的作息运转。关于失眠陷阱的解法,见:Curing DSPS and insomnia。
如果你无法重置相位,并在规律早起时仍持续感到疲倦,请考虑选择一份对你的身体可接受的工作,而不是反过来。你的长期健康和福祉正受到威胁。如果你绝对无法不靠闹钟生活,至少可以从改变你对睡眠重要性的心态开始,并确保不把坏习惯强加给孩子。也许年轻一代足够幸运,能在弹性工作制下工作,从而获得充足的不被打扰的睡眠。至少,不要树立坏榜样!
叫醒总统(Wake up the President)
2000 年 4 月 22 日夜间,在对小古巴流亡者 Elian Gonzales 迈阿密亲属住所那场臭名昭著的移民局突袭之前,比尔·克林顿总统被电话叫醒两次。他大概是历史上被最频繁打扰、睡眠最被剥夺的总统。直到心脏手术后,克林顿才开始认真对待饮食、睡眠和(真正的)运动。那些被打断的夜晚,肯定会影响他的表现和决策质量!有没有人想过一条规则:不要叫醒总统? 一条只有在真正的国家紧急状态下才能被撤销的规则?小布什总统(生于 1946 年)在 2001 年一架美国间谍机降落中国时被叫醒,2002 年耶路撒冷发生自杀式爆炸后也被叫醒。老布什(生于 1924 年)和希拉里·克林顿把”半夜被叫醒”作为他们竞选总统的资本和能力的一部分。似乎只有罗纳德·里根对自己睡眠的保护有相当严格的规定。他也以内阁会议期间打盹而闻名。他睡过了几次国际事件,而对他略显迟缓的决策没有明显的负面影响。批评者会说他睡过了整个伊朗门事件(Iran-Contra affair)。里根如此保护睡眠,是因为他比其他总统更懂睡眠的作用,还是他只是比别人懒一点?我不知道。不过,他确实树立了一个好榜样。
闹钟怪物(Alarm clock monsters)
Andrea K. 带着怀疑写信给我:“拿走一个普通人闹钟,他不会在想要的时间自己醒来,他们会错过工作、学校或其他什么。闹钟不可能那么糟,因为大多数人都在用,我从没注意到他们有什么问题:) 我家每个人从小就一直在用闹钟,没人突然发疯或开始变成怪物(至少目前还没有)!” 正如我之前写的,当你在清晨用闹钟去上班或上学时,你切断了睡眠的后几个阶段。如果对自然睡眠的侵入不大(例如几分钟到一小时),损害可能有限。如果闹钟切到夜间睡眠的中段,它的危害会大得多。你可以把使用闹钟比作吸烟或吃热狗。危害没有大到立刻被察觉的程度。公众花了多年时间才在很大程度上接受”吸烟有害”或”快餐有害”。很难量化损害的程度。然而,随着我们迈向知识社会,我们的智力表现变得越来越重要,睡眠剥夺的影响将受到更密切的审视,闹钟注定会逐渐失宠。与热狗不同,它们已经被用户普遍痛恨。如果作息足够规律,大多数人能在一定程度上把睡眠适应到日程上。当这些人需要诉诸闹钟时,他们切断的睡眠较少,损害也相应较小。尽管如此,我们应始终致力于彻底消除闹钟。最重要的是,我们应保护孩子免受被打断的睡眠之苦!
睡眠惰性(Sleep inertia)
什么是睡眠惰性?(What is sleep inertia?)
睡眠惰性(sleep inertia)是睡眠后可能伴随的昏沉感。睡眠惰性有不同的类型,术语上存在巨大的混乱,科学文献中不同类型的睡眠惰性之间也有大量混淆。睡眠惰性的一个令人困惑的定义示例:“睡眠惰性指大多数人醒来后经历的昏沉感。” 一个更恰当的定义应是:“睡眠惰性指因睡眠被打断或其他违反睡眠卫生的行为所导致的昏沉感。” 最重要的是,睡眠惰性并非人类睡眠中不可避免的一部分。在健康个体中,睡眠惰性是睡眠艺术中错误的直接结果。只要一丝不苟地遵守睡眠卫生原则,你根本不必经历睡眠惰性及其对学习、注意力、健康等的负面影响。
所有对睡眠惰性的研究都应清楚区分其不同类型:
- 被打断的深睡眠(interrupted deep sleep)——在示例性的睡眠结构图(hypnogram)中,我们可以看到第 4 阶段 NREM 在入睡约 30 分钟后出现。在这个阶段叫醒受试者尤为困难。从深睡眠中被唤醒时,我们体验到一种压倒性的、想要回去继续睡的需求。这种感觉可以比作头部被重物击中。大脑处于一种完全不适合处理清醒信息的状态。这正是许多多相睡眠者通过试图打断在他们主观夜晚相位进行的”小睡”,而强加给自己的折磨。
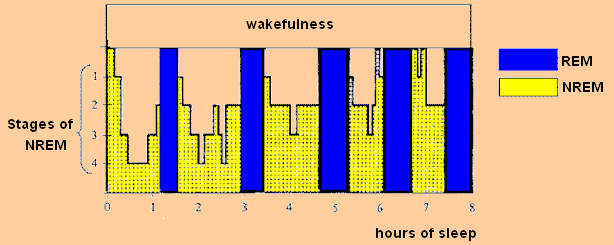
-
被打断的 REM 睡眠(interrupted REM sleep)——在睡眠结构图中,REM 睡眠以蓝色标出,其出现频率在夜间睡眠后期增加。打断完全展开的 REM 睡眠同样令人不快。然而,大脑的神经激素状态与打断深睡眠时完全不同。一个容易识别的特征是肌肉无力。由于 REM 的功能,运动系统在这个阶段被关闭。如果你在手握力计上测试力量,你得分可能还不如你的孩子。被打断的 REM 也常与梦境回忆相关。一旦大脑回到典型的清醒模式,REM 梦境记忆消散得非常快。如果你不立即写下梦境,5 分钟后它就可能从记忆中无法恢复。
-
在错误的相位醒来(waking in a wrong phase)——即便是自然醒来也可能导致睡眠惰性。如果你在主观夜间睡眠期间自然醒来,可能会在一段时间内感到相当昏沉。过早醒来的主要原因是过早的就寝时间。在较早相位睡眠的情况相当普遍。许多人不得不违背生物钟早起。他们常使用各种手段来早睡(从酒精到安眠药)。结果,他们早睡、早醒,看似获得了”充足”的睡眠却仍感到疲倦、没有精神。这是因为他们醒来时,身体在昼夜节律上仍处于主观夜晚期间。过早的就寝时间常导致失眠。然而,当睡眠成功启动时,睡眠控制系统可以提前启动一次相当于夜间睡眠的过程。这种睡眠可能走完全程,甚至持续稍长一些。然而,其结构会有所不同,而且早晨的昼夜节律睡眠倾向在醒来时仍不会被完全清除。结果,早晨的昏沉感作为昼夜节律困倦和各种由睡眠结构变化(例如 REM 亏欠)引起的睡眠亏欠的组合而产生。由过早醒来引起的惰性比打断 NREM 或 REM 睡眠更容易对抗,因为它在很大程度上随昼夜节律睡眠倾向的到期而消散。然而,轻微的睡眠变量亏欠可能持续整天。对某些人来说,在错误相位睡眠是他们日常生活如此司空见惯的一部分,以至于他们往往忘记什么是敏锐警觉的头脑,并且,如上面引用的错误定义那样,他们倾向于认为这是”大多数人经历的那种睡眠惰性”,仿佛它是正常人体生理的一部分。他们可能会用”我就是一直觉得累,这就是我”来打发它。从技术上讲,睡眠惰性的定义还应扩展到由就寝时间大幅延迟引起的大脑状态。这种大脑状态类似于过早醒来引起的惰性。它可能感觉更自然,因为它结合了睡眠倾向的两个组分:内稳态和昼夜节律困倦。它感觉不像惰性,而像是可能接近恶心的严重困倦。这种惰性对驾驶员尤其危险,因为它只会随时间恶化,并可能导致部分大脑皮层简单地进入睡眠模式(脑电图中出现 theta 波)的微睡眠时刻。
睡眠惰性是否表现出昼夜节律?(Does sleep inertia show a circadian rhythm?)
这个问题没有直接的答案。无论你读到什么关于这个主题的内容,都要确保把包罗万象的术语”睡眠惰性”拆解开来,对每种类型的睡眠惰性分别问同样的问题。如果你打断了深睡眠,它总感觉很糟。那种感觉的程度可能取决于睡眠的深度、你的内稳态状态,以及在较小程度上你的昼夜节律状态(只是因为深睡眠在很大程度上是内稳态性的)。然而,如果你打断 REM 睡眠,它更可能在昼夜节律 REM 峰时刻产生更深刻的影响。最后,错误相位的惰性纯粹是昼夜节律性的。它只会在你主观夜晚期间袭击你,并在你主观白天时自行消散。
如何从睡眠惰性中恢复?(How can I recover from sleep inertia?)
你可以在网上搜出数十种对抗睡眠惰性的疗法(示例),你可能会惊讶于所有这些”网络建议”背后的推理存在一个巨大的漏洞,它们往往没有注意到:时机恰当的睡眠是对抗所有形式睡眠惰性的最佳疗法!
对于被打断的睡眠惰性(无论是 NREM 还是 REM),简单的解药是:回去继续睡。惰性越强,你快速重新入睡的机会越大。像咖啡或运动这样的疗法可能让你感觉好些(也可能不会),但它们可能造成自己的损害。如果你的职业要求你在半夜醒来,请记住,你是在以自己的健康和长寿为代价履行职责。
错误相位的惰性更难对抗。在很多情况下你将无法重新入睡。更糟的是,试图睡觉有时会让事情更糟。最好的解药是熬过不适,避免在你下一个主观夜晚时段之前小睡,并在正确的相位入睡。大多数时候,足够长的清醒和命中正确相位会让你瞬间同步所有睡眠变量。然而,在某些情况下,昼夜节律的涟漪可能拖延数天,尤其是如果你不太擅长计算自己正确的睡眠相位。如果你做大量轮班工作或洲际飞行,很容易对自己主观夜晚何时到来感到困惑。在这种情况下,你可以使用 SleepChart 免费版来获得一些让猜测更容易的可视化支持。
睡眠会损害学习吗?(Can sleep impair learning?)
令人惊讶的是,对睡眠惰性类型的混淆还催生了另一个迷思:学习之前的睡眠会增加遗忘!时机恰当的睡眠不会引起睡眠惰性,也不会导致学习下降。恰恰相反,自然醒来后 20-60 分钟是学习效果最好的时候。当然,这仅在自由睡眠中成立。闹钟常常被打断夜间睡眠,而清晨相当不利于学习。
为什么小睡会引起睡眠惰性?(Why naps cause sleep inertia?)
只有当小睡在以下情况下进行时,才会引起睡眠惰性:
- 太晚,或
- 在严重睡眠剥夺的条件下,或
- 在 REM 睡眠亏欠的条件下。
这三种情况都能欺骗睡眠控制系统,让它以为这次小睡是启动一整夜睡眠事件的适当时机。如果在远早于主昼夜节律低谷(夜间顶点)之前试图启动完全展开的睡眠,你可能会过早醒来,并感觉获得了一次不完整、不令人神清气爽的夜间睡眠。这种睡眠会让你昏沉,并使你在主观夜晚期间更难启动恰当的睡眠。因此,要避免与小睡相关的睡眠惰性,首先要避免睡眠剥夺,并阅读关于最佳小睡时间窗的内容。
长睡眠与睡眠惰性(Long sleep and sleep inertia)
许多人认为长睡眠会导致睡眠惰性、头痛等。长睡眠后问题的根本原因是先前的睡眠剥夺或在错误相位睡眠。在自由睡眠作息下,健康个体不可能有异常长的睡眠。通常正是严重的睡眠剥夺,使得在大早于最佳昼夜节律入睡时间之前入睡成为可能。这种异常长的睡眠会贯穿主观傍晚和整个主观夜晚,累积到一些非常异常的睡眠总量(12-18 小时)。这种睡眠之后常出现一种类似睡眠惰性的状态(“磨损”综合征)。难怪人们很容易在长睡眠与睡眠惰性之间建立错误的联想。坚持长睡眠作息非常困难,因为睡眠调节机制会很快把睡眠时长带回更典型的范围。一方面,如果睡眠时段相对于昼夜节律周期调整不当,“磨损”综合征似乎可能持续。另一方面,“磨损”的观察通常来自那些工作日睡不够、周末睡得长的人。在后一种情况下,由于下一周的义务,后续观察常常无法进行。这加深了”睡得太多有害”这一错误信念。健康个体不可能”睡得过多”! 他们的大脑只会在正确的时间产生自然醒来。Jim Horne 和 Daniel Kripke 博士可能会另有说法。也许他们从未在峰值警觉窗口试着打盹?
轮班工作和时差的健康影响(Health effects of shift-work and jetlag)
工业化国家近 20% 的人口从事轮班工作!调查显示,只有 10% 的轮班工作者对他们睡眠作息对健康和生活的负面影响没有抱怨12。如果轮班设计得当,这些数字会好得多。这自然不会改变所有形式的睡眠调节都有风险且可能不健康的事实。研究表明,轮班工作者患有各种胃肠道和心血管问题。心血管变化可能由诸如 C 反应蛋白等炎症标志物介导。许多人难以获得令人神清气爽的睡眠。经过多天的慢性睡眠限制后,会累积显著的认知下降,这种下降最终接近严重急性完全睡眠剥夺的水平。轮班工作者中的物质滥用也远高于平均水平。看似轻微的问题,如头痛、注意力不集中、性欲下降、疲劳、易怒等,加起来对绝大多数作息设计糟糕的轮班工作者来说构成了相当悲惨的生活。困扰轮班工作者的那组问题,对研究时差的研究人员来说非常熟悉。已经为这两组相关症状分别创造了医学术语:轮班工作障碍(shift work disorder,SWD)和时差障碍(jet lag disorder,JLD)。关于时差最戏剧性的发现是,从事较长时期洲际飞行工作的空乘人员,其海马体出现细胞丢失(Cho,200113;Cho 等,200014)。我们可以推测,完全相同(放大十倍)的健康问题,会困扰那些能坚持多相睡眠作息足够长时间的人。
除了睡眠相位错位的直接影响外,轮班工作和时差还存在一定程度的睡眠剥夺。充足的睡眠对正常的葡萄糖代谢和预防肥胖及2 型糖尿病很重要。睡眠限制会降低瘦素(leptin)水平,并对饥饿素(ghrelin)产生相反的影响。结果,这两种食欲激素使睡眠剥夺的个体比休息良好的人感觉更饿,并上调了热量平衡稳态中体脂重量的设定点。每晚把睡眠限制在 5 小时,会导致这些食欲控制激素水平约 20% 的变化。这一变化相当于在自由进食下额外摄入约 1000 千卡,或按能量计算每月超过 3 公斤脂肪。睡眠限制可以轻易让胰岛素敏感性减半,导致2 型糖尿病,并显著增加高血压、中风、心脏病发作或肾衰竭的风险(Van Cauter 等,200715)。其他激素变化包括甲状腺激素水平升高(Allan 和 Czeisler,199416)、催乳素、LH和雌二醇(Baumgartner 等,199317)。最后,许多相位移动问题的根本原因,是轮班工作和时差对压力激素皮质醇昼夜水平变化的复杂影响。皮质醇水平变化影响的净效应,是长期来看导致身体”自我消耗”的过度分解代谢状态。这样,忽视你的生物钟,你会同时变得肥胖和生物学上”被消耗殆尽”。
2007 年,国际癌症研究机构(IARC)发表声明称:“涉及昼夜节律破坏的轮班工作对人类可能致癌。” 使用”致癌”一词可能略有误导,因为轮班工作者癌症增加的实际原因,可能与免疫功能下降及身体对抗突变癌细胞自然能力下降有关。然而,这一声明很重要,因为它为轮班工作和时差的命运定了性——它们最终应归入那些造成广泛而严重的系统性健康问题的长期健康风险因素类别。
设计糟糕的轮班工作、时差和睡眠剥夺,都是影响你福祉和长寿的严重系统性健康风险。
关于生物钟这个微小而精致结构的更多内容,见专门介绍视交叉上核(SCN)的章节。
设计得当的轮班工作(Properly designed shift work)
我常把轮班工作限定为”设计糟糕”的健康风险。这是因为可以为一群人设计出让昼夜节律破坏最小的作息。使用时间疗法(chronotherapy),可以逐步把员工调入通宵工作。这种疗法的主要原则是:相位移动每天不应超过一小时,并且除少数例外应是向前移动(即一天长于 24 小时,而非更短)。任何偏离这一原则、并涉及休闲时间、小睡、强光、褪黑素、安眠药、莫达非尼等的疗法,都是纯粹浪费时间,因为它们一直在对抗不可避免的事:工作时间与主观夜晚时段之间的错位。这种错位只能通过逐步的、时机恰当的、基于相位移动的调整来补救。
尽管许多轮班工作者会不同意我(主要出于心理和便利原因),我坚持认为,长时间维持夜班(例如一个月)比在身体时钟没有任何调整机会的情况下在夜班和白班之间规律轮换,要更容易、更健康。一些癌症研究者也反对长时间上夜班,因为有记录的褪黑素下降,而褪黑素被认为具有抗癌特性1819。然而,这些需要与昼夜节律破坏这一更严重的问题权衡考虑。
过度睡眠(Excessive sleeping)
关于睡眠最持久的迷思之一是,我们的身体被设定为尽可能多地获取睡眠。甚至一些有声誉的研究者也认同这一观点。他们把睡眠比作过度进食。有人指出因纽特人冬天睡得多长,另一些人指出,被允许自由睡眠的人常常严重暴睡,累积不光彩的睡眠时长,仿佛节能是睡眠的主要功能,仿佛所有动物都像永远饥饿一样懒惰。
一些科学家甚至设想与热量限制类似的睡眠限制。可以想象,睡眠限制在某些罕见病例中对病人可能有益(例如”醒来吃药”)。然而,它与热量限制的类比,与反向提议——清醒限制——一样站不住脚。这个迷思可能源自流行病学研究,这些研究显示每晚睡 7 小时的人比睡 9 小时的人活得更长。然而,建议通过限制睡眠来延长寿命,就和因为非常矮或非常高的人不如街上普通人活得长,就试图把人缩小或拉长一样”聪明”。
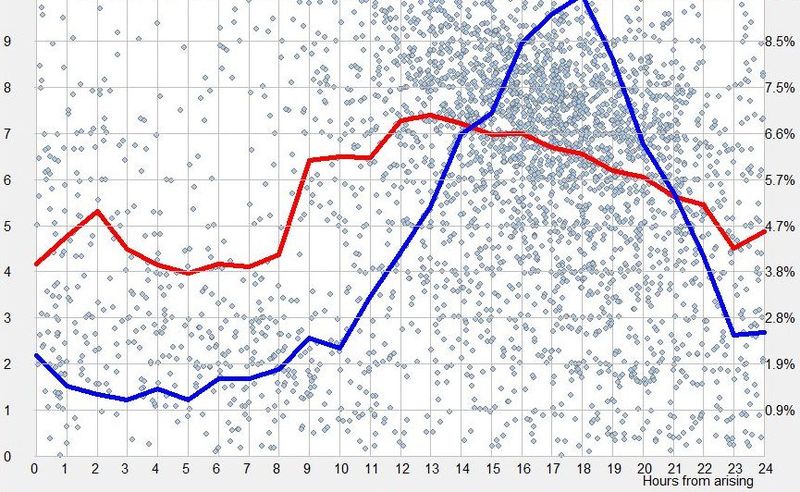
我们无法证明,获得超过神经所需睡眠有任何演化优势。过度睡眠的有害迷思可能让你认为,自由睡眠会让你睡得更长,就像自由进入厨房会让你过度进食一样。考虑到已知的睡眠功能,超过标准的 6-8 小时睡眠没有特定益处。睡眠是清醒期间积累的好处(如学习、运动等)的神经生理消费者。其健康的内稳态和昼夜节律控制大致确保了睡眠与清醒的最佳比例。在自由睡眠条件下暴睡的人,通常来自长期的睡眠剥夺,或在相对于其昼夜节律相位过早的时候启动睡眠。他们的总睡眠时间在自由作息几天后很快降到自然平均水平。一项研究表明,要获得超过 8 小时的不间断睡眠,应在体温最低点(nadir)前约 6 小时(即警觉性顶点之后不久)启动睡眠(Dijk 和 Lockley,200220)。在 SleepChart 数据提交中也能看到同样的现象。例如,在所展示的图中,睡眠启动时间比最偏好的入睡时间早 3 小时(仅在傍晚”禁区”之后一小时)时,获得最长睡眠时长。这些观察彻底否定了我们有过度睡眠倾向的观点。
显示”过度睡眠”在睡眠启动过早时发生的昼夜节律图。在图中,第 16 小时启动的睡眠长于平均水平,而睡眠-清醒周期不等于 24 小时(相位前移的不平衡周期)。相比之下,在偏好的第 19 小时启动的睡眠短了近一小时,并产生完美平衡的 24 小时睡眠-清醒周期。
如果你主要关心的是时间,你可以少睡一些,以脑力敏锐度为代价获得更多时间。如果你主要关心的是脑力,你应遵循这句格言:睡眠的最大效率,是在没有人为睡眠调节(即没有闹钟、药片、设计师作息、物质等)的情况下睡眠时实现的。自由睡眠作息会让你平均睡得更少。它会让你比任何迫使你偿还累积睡眠债的人为睡眠作息睡得少得多。不规律的作息注定会产生亏欠,因为你只能通过干预才能实现不规律的睡眠。关于过度睡眠的更多阅读,见:Jim Horne and Daniel Kripke。
睡眠习惯(Sleep habits)
在本节中,我想说明人们的睡眠习惯可以差异巨大,而其中一些差异有着重要的生物学根源。科学家用睡眠时型(chronotype)一词来区分不同个体的睡眠时间和时长偏好。一个人的时型可能使他成为短睡眠者;另一个人的时型会让医生诊断为睡眠相位障碍。尽管表面上千差万别,少数几个底层变量应该能让你相当容易地搞清楚自己的时型。你的时型可能决定你是否适合某些职业。幸运的是,你不必在选择专业或工作前就确定自己的时型。许多人会自然而然地被与自己睡眠习惯匹配的活动和职业所吸引。医生或消防员需要耐受轮班工作和被打断的睡眠,送奶工起得早,而健身房或迪斯科舞厅老板需要熬夜,作家则可以是任何时型,因为他/她可以把写作时间适应到自己的睡眠模式。为了说明个体睡眠模式,我使用一个名为 SleepChart 的免费程序(Wozniak 等,200321),你可以在此下载来可视化地绘制自己的睡眠。如果你收集了几个月的数据,我将非常乐意收到你的数据文件用于分析和未来研究。从 SleepChart 发送数据只需在程序中点击一次。
生物钟(Body clock)
睡眠与清醒的循环由生物钟调节。生物钟位于大脑中,主要位于视交叉上核(suprachiasmatic nucleus,见专门介绍 SCN 的章节)。生物钟的周期约为 24 小时。在一个 24 小时的一天中,我们有 5-10 小时非常困倦,这正是我们通常睡眠的时间。在其余的 14-19 小时里,我们通常保持清醒,或在午休时间小睡。如前所述,只有一小部分清醒时间适合最高质量的脑力付出(见:Optimizing the timing of brainwork)。最大警觉性的时段可能短至 2-4 小时。我们应以这样的方式安排一天:让睡眠出现在最大困倦的时刻,而那些需要最大专注或创造力的活动落在最大警觉性的时段。强迫身体和生物钟改变清醒活动和睡眠的时机,非常困难且通常非常不健康。反过来则容易得多:让自己的生活适应由生物钟支配的自然周期。这种适应取决于一个人自身生物钟的独特属性。在以下章节中,我将试图展示由给定个体的生物钟属性所决定的不同睡眠习惯类型。
相位障碍中的睡眠组分(Components of sleep in phase disorders)
调节困倦感的主要有两个机制(见:Two components of sleep)。一个是生物钟,另一个是”清醒计量器(wake-meter)“。生物钟每 24 小时产生一次困倦感的增加。清醒计量器则随清醒时间的延长而增加困倦感(即我们清醒得越久,越困)。在睡眠文献中,这两个机制被称为睡眠倾向的昼夜节律组分和内稳态组分。
睡眠控制组分:
- 昼夜节律时钟(circadian clock)——以 24 小时为周期产生困倦感。
- 内稳态控制(homeostatic control)——清醒计量器测量我们保持清醒的时间,并在我们清醒足够久后触发困倦感。
在受 DSPS 或 ASPS 影响的人中,可能存在若干因素的组合,使得在正常时段获得好睡眠更加困难:
- 昼夜节律时钟运行的周期远不同于 24 小时。例如,DSPS 患者的时钟可能被设定在 25-26 小时。
- 昼夜节律时钟对重置时间的因素(称为授时因子(zeitgebers))不敏感。正常人通过清晨的光照和活动重置时钟,此外傍晚的黑暗和不活动也为时钟提供线索。具有正常生活方式的正常人能轻易地把睡眠与日夜周期同步。
- 内稳态清醒计量器有异常的时间常数。敏感的清醒计量器会让人在醒来后很快感到疲倦;不敏感的清醒计量器可能让人倾向于长时间保持清醒。咖啡因滥用可能通过腺苷受体下调导致警觉性快速下降。
- 生活方式对昼夜节律和内稳态睡眠调节机制的行为有巨大影响。同一个体在不同因素下会显示不同的睡眠模式,这些因素包括:使用人工照明、运动、压力水平、令人兴奋活动的时机、小睡、饮食、气候、环境温度变化、健康状况等。
百灵鸟与猫头鹰的误解(Lark-owl misconception)
研究表明,15% 的人会把自己归为”早晨型”即百灵鸟(lark)。另有 20% 会自称为”夜晚型”即猫头鹰(owl)。其余 65% 是 indifferent 或”中段型(mid-range)“。你是哪种类型?你可以在网上找到许多百灵鸟或猫头鹰测试。然而,我至今没见过哪怕一个设计良好、能真正回答你基因倾向问题的测试。尤其是,同一个人在工作日作息下可能被归为一种时型,而在自由睡眠作息下又被归为另一种。
很少有人知道,他们可以通过时间疗法(例如每天把睡眠时间推迟 30-45 分钟)轻松地适应一个完全不同的作息。如果你让一个典型的猫头鹰每天晚睡 30-45 分钟,猫头鹰会不断把入睡时间推向更晚。起初它会在白天睡觉,然后睡眠逐渐移到更晚的时段,直到猫头鹰发现自己要在傍晚早早就寝,赶在百灵鸟之前起床!令人惊讶的是,即便最坚定的猫头鹰也能在那之后相当长时间舒适地坚持早起的时段!关于一天中的睡眠时间,几乎没有什么天然的偏好!
然而,有一个因素驱使人们相信自己是某种睡眠时间偏好类型。这就是昼夜节律周期的长度及其驯化到 24 小时的能力。如前所述,典型的昼夜节律时钟周期长于 24 小时。周期特别长的人倾向于每天越来越晚入睡,把清晨时段的极限推到强制起床时间导致无法忍受的困倦为止。换言之,周期长的人只要有可能,就倾向于在夜间工作、在早晨睡觉。
百灵鸟和猫头鹰在偏好的睡眠时间(相对于白天)上并无不同!差异来自昼夜节律周期的长度、对授时因子的敏感度以及生活方式。你可以轻松地让一只百灵鸟舒适地工作到深夜,也可以让一只猫头鹰在凌晨 3 点起床——这可以通过时间疗法(周期调整)做到!此外,如果猫头鹰采纳古老农民的生活方式(例如放弃用电),它们也能一直坚持在黎明起床。
较小比例的人会经历短的昼夜节律周期,并在傍晚早早就感到极度困倦。这就是百灵鸟型。生活迫使百灵鸟比其自然偏好稍晚入睡(家庭、工作、光照等),这使百灵鸟保持与时间同步,它们常会声称清晨的宁静、鸟鸣或日出的美景让它们坚持早起。然而,仍然可以强行推动一只百灵鸟逐渐移动睡眠时间,表现得像一只猫头鹰!
在现代社会,只有一小部分人能夸耀拥有完美稳定、规律的自然睡眠模式。这些人不仅是最健康的人,也是在睡眠和清醒仪式方面最有规律的人。他们一丝不苟地遵守自己的仪式,避免使用闹钟,避免晚间娱乐,避免影响睡眠的药物等。与这些调节良好的人不同,被移到早晨作息的猫头鹰会逐渐倾向于回到其标准的深夜节律。同样,百灵鸟会很快回到与鸟儿一同起床。
一些相关研究表明,猫头鹰(以褪黑素释放的时机定义)比百灵鸟表现出略高的智商(Roberts 和 Kyllonen,199922)。
用 SleepChart 绘制睡眠(Charting sleep with SleepChart)
理解产生睡眠和清醒的控制机制,对理解睡眠习惯极有帮助。这对受多种睡眠障碍(尤其是失眠和相位移动障碍)困扰的个体尤为有用。对昼夜节律变量的简单测量和简单的时间疗法工具,可能为那些常常与失眠、令人不满意的睡眠、或在错误时段睡眠斗争多年的人带来健康的睡眠。更好地理解时间生物学,也有助于消除危险的做法,如计划不周的轮班工作、对时差健康后果的无视、累积的睡眠剥夺,以及网络上的 Uberman 睡眠风潮。
为了说明各种睡眠习惯,我使用一个名为 SleepChart 的免费程序的图表。你可以在此下载 SleepChart,并从今天开始你自己的分析。你只需在程序中点击睡眠区块的开始和结束即可。查看 SleepChart 窗口底部,了解鼠标指针对应的确切时间。如果你设置了错误的区块,点击选中它并按 Del。
使用 SleepChart 数据,我将试图解释健康人可能无法获得令人神清气爽睡眠的主要原因:睡眠相位问题(sleep phase problems)。
SleepChart 试图近似与最大困倦感、低体温、低 ACTH、高褪黑素等相关的昼夜节律顶点(acrophase)。其基本假设是,当你用 SleepChart 记录睡眠时,你没有试图人为地操纵睡眠时段。每一次对睡眠作息的干预都会使 SleepChart 工具的准确性下降。以下是最应避免的干预:
- 用闹钟醒来,
- 在傍晚与困意对抗(例如为了推迟睡眠),以及
- 用物质控制睡眠(例如酒精、安眠药等)。
在你偶尔推迟睡眠或使用闹钟的罕见情况下,你可以用相应的标记取消该睡眠事件的资格。然而,所有修改睡眠作息的尝试都会在一定程度上欺骗算法,你的读数会不准确或完全错误。同样非常重要的是,在确定你最佳睡眠时段时,你不应试图遵循昼夜节律近似值!你应始终优先考虑你身体的自然信号,即困倦感。遵循 SleepChart 的近似值可能导致错误的正反馈。换言之,图表中的错误可能被你试图遵循图表的行为放大,从而扰乱睡眠周期。最坏的情况下,你甚至可能在没有真正患该障碍的情况下自我诊断为 DSPS!你唯一且仅有的”去睡觉”标准应是迅速增加的困倦感。你可以用图表来近似计算准备就绪入睡的时刻,以便你能及时”冷却”下来。你也可以发现 SleepChart 在 ASPS 或 DSPS 的时间疗法中有帮助,让你更容易地在不与自然睡眠节律冲突的情况下安排日程。
得益于众多贡献者提交的 SleepChart 数据,我们可以得出一些有趣的结论。最令人信服的大概是这一假设的证实:我们可能正面临**年轻一代延迟睡眠相位综合征(DSPS)的流行,尤其是在学生和从事高科技工作的人群中。这一流行病是人类及其灵长类祖先在过去几百万年中所处环境,与我们今天所处的环境(电照明、互联网、电脑、电视、激烈竞争和 24 小时社会)之间日益扩大的差距的结果。生活方式与生物学之间日益扩大的鸿沟,驱使许多人寻求激进的解决方案并采取极端措施。对那些尝试适应 Uberman 睡眠作息的人的快速调查揭示了一个有趣的真相:尽管把更多清醒时间塞进一天的想法很有吸引力,但大多数”实验者”开始对多相睡眠产生兴趣,正是因为难以获得令人神清气爽的睡眠!
一些人对把 SleepChart 作为睡眠辅助工具的概念持怀疑态度:“这太复杂了,奥卡姆剃刀需要刮一刮!睡眠就像呼吸空气或喝水一样自然,如果你必须设置复杂的图表和实验,以及完全古怪的睡眠-活动模式才能获得像样的睡眠,那你一定有问题——但更多是心理而非生理的。” 确实,睡眠在自然环境下会自然发生。麻烦始于我们用咖啡因、酒精、尼古丁、人工照明、24/7 社会、夜间娱乐等干预自然。SleepChart 看似复杂,但对那些可能有睡眠问题的人来说,它可能仍是预测自由睡眠条件下最佳睡眠时机的最简单方法。SleepChart 只会问你何时入睡、何时(自然)醒来。所有计算复杂度都隐藏在后台。近似过程无需用户进一步输入,它预测昼夜节律顶点和最佳入睡时间。SleepChart 甚至能解开睡眠的内稳态和昼夜节律组分。理解这些也有助于规划健康的睡眠。
我同意,诉诸 SleepChart 这类工具是麻烦时代的标志。然而,SleepChart 有良好的记录,能帮助人们理解他们看似不规律的睡眠模式并组织他们的睡眠。入睡可能很自然,但有许多因素掩盖困倦感或放大它。对作息极不规律的人来说,这可能构成难以逾越的障碍!
有睡眠问题的人常常被那些”天生好眠者”误解:“如果你每天扎扎实实工作 8 小时,吃 3 顿像样的饭,有正常的家庭生活,把别人当人看待,那么晚上你会被幸福地击倒上床,第二天早上幸福地神清气爽。当然,历史上大多数人都一直如此,当然也将永远如此。” 这种对睡眠问题的态度,与告诉一个临床抑郁症患者”振作起来”没有两样,或指望一个海洛因成瘾者突然戒断并立即恢复正常生活。一个备受折磨的失眠症患者,如果被告知失眠来自”工作不扎实”、“饭吃得不体面”、“家庭生活不正常”或”把别人不当人看”,只会对自己更加沮丧。麻烦源于生物学与现代生活方式的冲突。随着人工照明的到来,睡眠障碍统计数据飙升,并因电视、电脑游戏和互联网而雪上加霜。随着移动电话和即时通讯的出现,失眠和睡眠相位障碍似乎达到了流行病的程度。越来越少的人能够把工作抛在脑后、应对压力或放弃晚间活动。在没有生活方式重大改变或昼夜节律控制方法突破的情况下,受生活方式相关睡眠障碍困扰的人面临着在每日睡眠剥夺的痛苦与扔掉闹钟这样的激进解决方案之间的选择。当然,我们可以期待科学给出答案。但在那之前,“幸福地神清气爽”地醒来,仍是工业化国家中日益缩小的少数人的特权。
SuperMemo 中的 SleepChart(SleepChart in SuperMemo)
为了分析睡眠与学习之间的联系,SleepChart 已被集成到 SuperMemo 快速学习软件中。与其解释 SleepChart 本身,我将简要描述它在 SuperMemo 中的功能。请注意,由于免费版无法访问你的学习数据,其中一些与记忆相关的功能未包含。
几年前,基于睡眠对学习至关重要的认识,SleepChart 被纳入 SuperMemo。要睡好、学好,需要理解自己的昼夜节律。SuperMemo 中的 SleepChart 正是为了协助这项任务而设计的。它可以帮助你优化睡眠时机以及学习时机。此外,你可以提交你的睡眠和学习数据用于分析,并在我们关于睡眠对记忆影响的研究中做出自己的贡献。你可以在 SuperMemo 中通过以下方式访问 SleepChart:(1) 主菜单上的 Tools : Sleep Chart,(2) SuperMemo commander,或 (3) 直接按 F12。
睡眠区块以蓝色标记,学习区块以红色标记。各天的总学习时间显示在右侧。选中的睡眠区块以黄色显示,其长度显示在底部。
在 SuperMemo 中,学习时间线是自动生成的。每次你用 SuperMemo 进行复习,学习区块就被添加到时间线(图中以红色显示)。另一方面,你的睡眠数据必须手动记录(以蓝色显示)。在最低技能水平,你可以用 SleepChart 对你偏好的学习和睡眠时段进行基本的视觉检查。然而,更高级的功能(如优化学习时机或睡眠时机)需要高级的分析和对昼夜节律的理解。如果你从今天开始记录睡眠数据,你将能够使用未来更高级的 SuperMemo 版本来研究和理解你的睡眠与学习。
SleepChart 中的睡眠时间线(Sleep timeline in SleepChart)
SleepChart 中的睡眠时间线必须手动记录。要记录一个睡眠区块,点击区块的开始(入睡)然后点击结束(醒来),你也可以先点结束。超过 22 小时的睡眠区块不被允许。睡眠区块不能与复习时间线重叠(你不能一边用 SuperMemo 学习一边睡觉)。如果你已经在 SleepChart 免费版中收集了睡眠数据,你可以通过 File : Import : SleepChart file 把数据导入 SuperMemo(也可以从电子表格导入)。如果你从 SleepChart 免费版导入文件,可以用 File : Verify : Block overlaps 检查睡眠与学习的重叠。防止区块重叠是在 SuperMemo 中使用 SleepChart 相对于独立 SleepChart 的一个重要优势——在独立 SleepChart 中,很容易在记录数据时失相位(例如漏填一天,一个月后才注意到)。你可以标记被强行推迟的睡眠区块,以及被闹钟或其他因素缩短的区块。请注意,如果你不人为地调节睡眠(如闹钟、安眠药等),你将获得最佳的分析结果。所应用的模型不会完全考虑人为干预。最后但同样重要的是,自然睡眠是你在学习和为最大健康与福祉方面都应追求的。
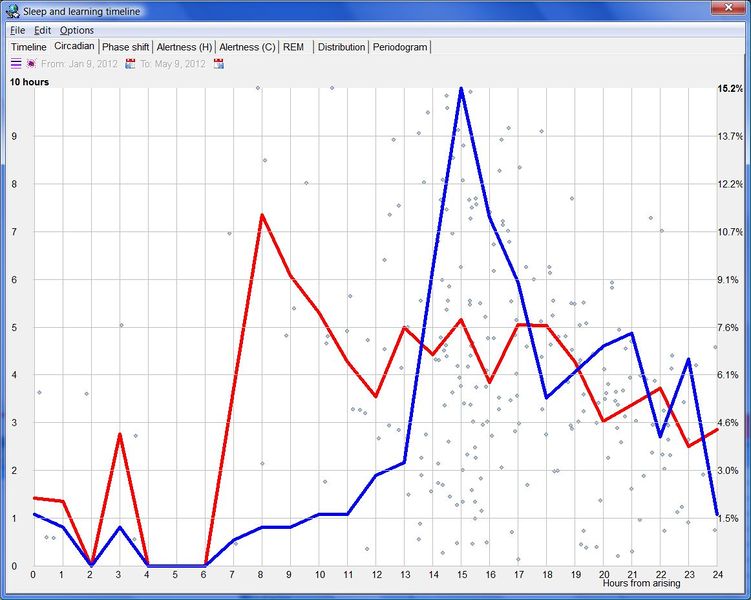
SuperMemo 中的睡眠与学习时间线(Sleep and learning timeline in SuperMemo)
把睡眠时间线与取自 SuperMemo 的复习数据结合,开启了一系列新的研究和优化选项。
与各天相关的各种睡眠统计数据显示在右侧。睡眠区块可以用工具栏上的 Consolidate 按钮合并。例如,如果你在夜里醒了 5-10 分钟,合并会让 SuperMemo 把整夜睡眠事件视为单一实体。短暂的夜间醒来是正常的,即便我们并未察觉,且对学习影响很小。睡眠区块合并常能揭示睡眠的重要属性(例如见:Preference for night sleep)。它有助于把连续的睡眠事件视为单一高睡眠倾向时期的表达。
除睡眠统计外,SleepChart 也可以估计最佳入睡时间。两个独立的模型用于预测夜间中点以及预计的最佳就寝和醒来时间。这些近似对那些轮班工作或因各种原因在不规律时段睡眠的人来说可能有用。例如,在一周不规律的睡眠之后,可能很难确定可能产生最佳睡眠质量的就寝时刻。入睡过早可能导致过早醒来(这常常毁掉整夜睡眠),入睡过晚可能导致夜间睡眠短、睡眠剥夺以及第二天警觉性下降。基于睡眠史预测最佳睡眠时间是一门不精确的科学,所用的两个模型可能产生不同结果。重要! 你的自然本能应始终优先于数学模型。此外,睡眠优化的最佳结果是在自由睡眠中实现的。如果你使用闹钟、强迫自己整夜清醒,或服用安眠药,模型可能无法充分考虑你睡眠控制系统中发生的混乱变化。
蓝色和红色连续线是使用 SleepChart 模型(基于睡眠统计)对最佳睡眠时间的预测。黄色连续线显示使用相位响应曲线模型对最大昼夜节律困倦(昼夜节律夜间峰值中点)的预测。注意,理论上黄线应大致落在蓝线和红线之间。然而,当睡眠模式的破坏严重时,这些线可能偏离,这表明很难构建出完全匹配受重大扰动影响的睡眠控制系统混乱行为的模型。青色点指向预测的白天警觉性低谷(即小睡可能最有生产力的时间)。
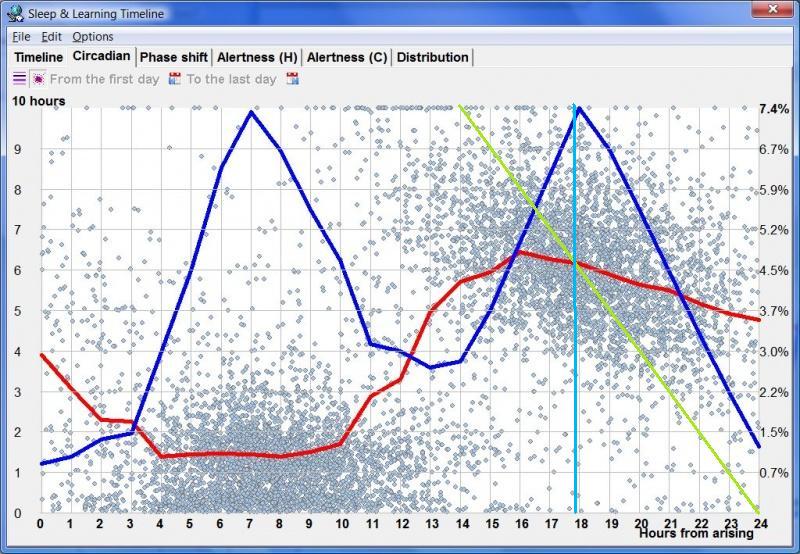
昼夜节律图(Circadian graph)
SleepChart 中的昼夜节律图可以帮助你更好地理解自己的睡眠模式,并可视化周期的不稳定程度(即你的睡眠-清醒周期有多难塞进 24 小时)。在图变得有意义之前,你需要几个月的数据。此外,睡眠日志中的主观夜晚近似线受显著的滞后(hysteresis)影响。如果你的生活方式发生剧烈变化(例如作为某种疗法的结果),近似线可能需要几周才能与数据正确对齐。此时昼夜节律图可能更难解读。在这种情况下,你可以使用 From the first day 和 To the last day 选项来划定感兴趣的时段,把分析限制在以某种生活方式为特征的选定时段。
单相睡眠者的示例昼夜节律图
睡眠启动(Sleep initiation)
蓝线显示偏好的入睡时间。它对应于从落入清醒日某小时的睡眠区块数量推导出的睡眠倾向,其中横轴的零点指醒来时刻。在任何给定时刻发起的睡眠事件百分比显示在右侧纵轴。蓝线大致表达你的”清醒带来的疲倦”,也表达你入睡的能力。你自己的最佳入睡时间是你个人特征,因人而异。对大多数人,最佳入睡时间落在醒来后 16-20 小时的范围内。在本例中,最偏好的入睡时间出现在清醒的第 18 小时。
睡眠维持(Sleep maintenance)
红线显示平均睡眠长度。这条线粗略反映了维持睡眠的能力,即最长的睡眠事件发生在主观夜晚。平均睡眠长度显示在左侧纵轴。该图会告诉你,即便你能在白天入睡,它也不会持续很久。在大多数规律睡眠者中,只有清醒 11-14 小时后,已启动睡眠的长度才开始增加。注意,睡眠长度图相对于偏好的睡眠启动时间略有相位偏移,这是因为长睡眠主要通过早入睡来实现。
优化入睡时间(Optimizing bedtime)
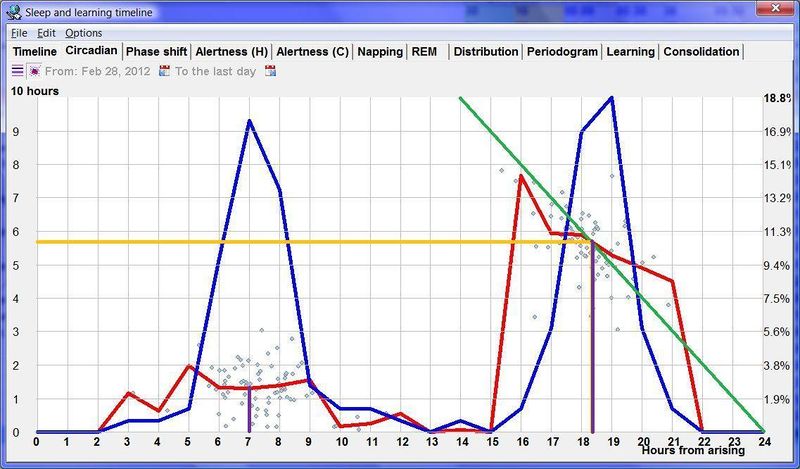
如果你试图确定最佳入睡时间,找到蓝曲线的傍晚峰值,并选择附近能产生足够长睡眠(红曲线足够高)的点。此外,注意你的清醒时间和睡眠时间加起来应等于 24 小时,否则你会经历相位移动。
有些人白天小睡,有些人则不睡。在小睡者中,蓝曲线也应指向最大的白天警觉性低谷。短的小睡时间实际上是良好小睡时机的标志,只要小睡不是相对于蓝曲线太早(见:Best nap timing)。不睡午觉的人也会在第 7 小时左右经历困倦峰值,尽管他们的蓝曲线不会显示为显著的凸起。
如果图表显示你的最佳小睡时间落在第 8 小时,而你早上 6 点醒来,你应在 14:00 左右休息一下,找一个僻静的地方休息几分钟。你也可以在 13:00-13:30 左右安排午餐,为午睡营造完美的条件。
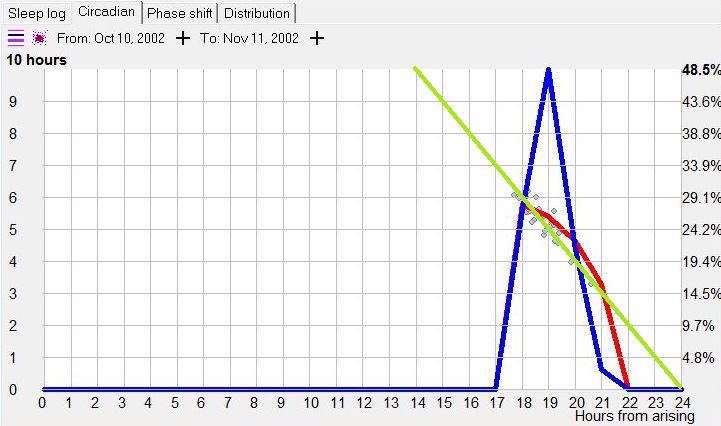
示例 1:不稳定的昼夜节律周期(Example 1: Unstable circadian cycle)
在下面的示例昼夜节律图中,平均而言,最佳夜间睡眠是在早晨醒来后 18 小时启动的(假设图表是在没有任何人为睡眠控制(如闹钟、推迟睡眠等)的情况下创建的)。蓝线显示第 18 小时是偏好的入睡时间,而睡眠长度(红线)足够长,加起来构成 24 小时的睡眠-清醒周期。
由于蓝色峰值高度相同,我们可以得出结论:该图代表一个一丝不苟的小睡者,其最佳午睡时间出现在醒来后 7 小时。在本例中,对于早上 8 点醒来,午睡应在下午 3 点开始,夜间睡眠在凌晨 2 点左右。对于两个蓝色峰值,7.4% 的睡眠事件处于最佳小时,而其余 85% 是次优的。
在第 16 小时可获得最大睡眠长度,然而这并不表明这是最佳入睡时间。如果睡眠启动过早,它可能赶上也可能赶不上主观夜晚的完整昼夜节律低谷。换言之,存在仅睡几分钟就过早醒来的风险。这样的醒来让人更难再次入睡。这是失眠的主要原因之一。再次启动睡眠的困难,源于睡眠过程中内稳态睡眠倾向的快速丧失。此外,在完整昼夜节律低谷之前启动的睡眠,似乎不比稍晚启动的稍短睡眠更有价值(例如,反映在早晨神清气爽的主观感受,或多导睡眠图的测量)。蓝色内稳态线表明,在第 18 小时更可能有效启动睡眠,其平均长度为 6 小时。如果你的图表是在不试图人为调节睡眠的情况下生成的,内稳态曲线中的第二个峰值通常指向最佳入睡时间。图还表明,如果睡眠推迟一小时,它将缩短 10-30 分钟。即便这一缩短也可能影响白天的表现。如果睡眠提前一小时,它可能长 10-30 分钟,但其质量不太可能按比例增加。
该图还可以显示昼夜节律周期的长度如何由入睡时间决定。绿线显示稳定的 24 小时睡眠-清醒周期的盈亏平衡点集,其中睡眠与清醒时间之和为 24 小时。绿线右侧的所有昼夜节律图点导致相位延迟,而另一侧的点将导致相位前移。青色线显示 24 小时周期绿线与红色睡眠长度线的交点。由于绿线和红线之间的夹角较大,这种睡眠模式相当不稳定。这意味着在第 18 小时之前入睡会导致周期短于 24 小时,而在第 18 小时之后入睡可能延长周期并导致相位移动延迟。例如,过早入睡(约第 15 小时)会导致一天持续 21 小时(横轴第 15 小时对应从红色曲线读出的平均睡眠长度 6 小时)。较晚入睡(约第 18 小时)会导致完美的 24 小时日,而非常长的清醒日(例如 20 小时)会产生持续 25.5 小时的一天。自然,所有对一天长度的操纵都应避免,因为过早入睡增加失眠的机会,而过晚入睡增加睡眠剥夺和 REM 睡眠亏欠的机会。理解自己的睡眠偏好对规划轮班工作或对抗长途飞行的时差非常有帮助。
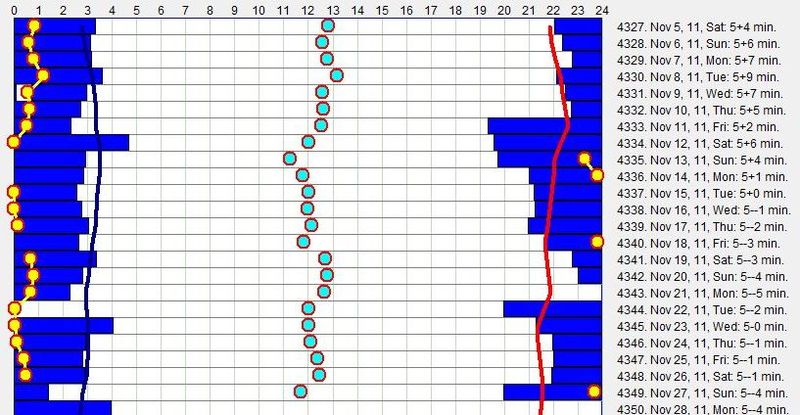
示例 2:稳定的昼夜节律周期(Example 2: Stable circadian cycle)
第二张图显示了一种比上面第一个例子稳定得多的睡眠模式。
该图显示了一个规律小睡者,偏好 19 小时的清醒日。与前面所示的图不同,由垂直青色线划界的稳定睡眠-清醒周期区域宽得多,因为红线和绿线几乎平行。这意味着如果睡眠在第 20 小时之后启动,夜间睡眠将被缩短以适应 24 小时周期。自然,即便推迟的睡眠不引起相位移动,它也总是因阶段压缩而导致较低的睡眠质量。这种睡眠会导致睡眠亏欠。持续少于 20 小时的日子可能导致相位前移。尽管自由运行,最长的平均睡眠时段(约第 16 小时启动)也不到 6 小时。这说明在自由睡眠中过度睡眠不是问题。在本图中,最佳午睡时间同样落在第 7 小时,且被一丝不苟地执行(超过 14% 的睡眠事件”准时”执行)。
相位移动障碍(Phase shift disorders)
如上面两张图所示,凭借足够的自律,有相位障碍的人应该能够实现 24 小时的自由运行节律,与所期望的醒来时间无关。在实践中,由于生活方式的各种扰动(考试、压力、社交等),以及需要早起的压力,对于有严重相位移动问题的人来说,遵守最佳的 24 小时睡眠作息可能非常困难。对于那些需要在特定早间时刻醒来的人,自由睡眠可能在没有闹钟、褪黑素或其他不受欢迎的措施的情况下无法实现。
24 小时睡眠周期(24-hour sleep cycle)
完美的 24 小时周期(Perfect 24-hour cycle)
现在让我们考虑一个理想同步的 24 小时周期。在下面的图中,一位八旬女性每天自然在凌晨 3:00-3:30 左右醒来。她每天睡 5.4-5.5 小时,醒来时神清气爽,全天活跃。
由于醒来时刻落在黑暗时段,与日光没有同步。该周期由傍晚活动同步,而非日光。受试者心中保持一个”必须睡觉”的时间估计,帮助生物钟与一天中的时间同步。这种”心理印记”在 2002 年 10 月 27 日周日夏令时结束后表现为睡眠节律的平滑变化(图表忽略夏令时,使得变化前的醒来时刻设定在凌晨 2:00-2:30)。
尽管据说衰老会增加夜间醒来(可能是由于睡眠控制中心的细胞丢失),但该受试者在研究期内报告没有醒来。
昼夜节律图显示第 19 个清醒小时有一个受偏好的入睡时间。由于平均夜间睡眠事件持续 5 小时,睡眠-清醒周期恰好持续 24 小时,且入睡时间的日常波动极小。由于绿色盈亏平衡线与红色昼夜节律线在 3 小时跨度内几乎平行,这种睡眠模式非常稳定,所有入睡时间的推迟都以睡眠时间为代价,而不引起相位延迟。
睡眠与压力(Sleep and stress)
压力可以破坏睡眠的肌理。下面的 SleepChart 图展示了压力对一个平衡良好的 24 小时睡眠模式的影响:
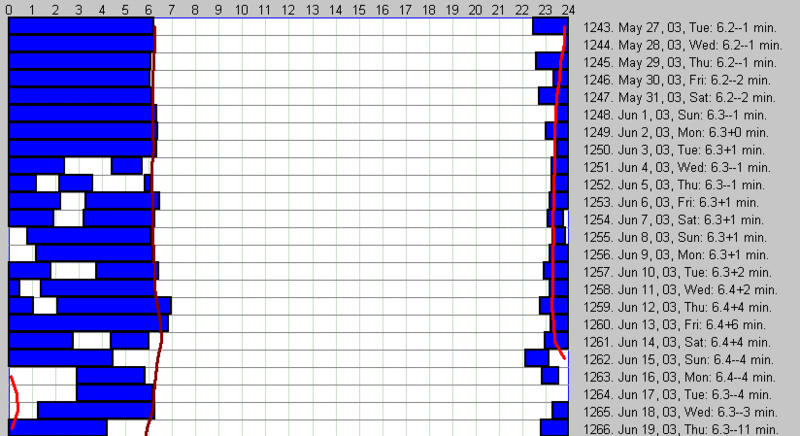
在所给的例子中,一位中年自雇男性每天自然在 6:00-6:20 左右醒来。然而,在 2003 年 6 月 3 日,一个严重的家庭问题把节律推入混乱,表现为频繁的夜间醒来。一个月后,随着家庭冲突的解决,节律回到了常态。
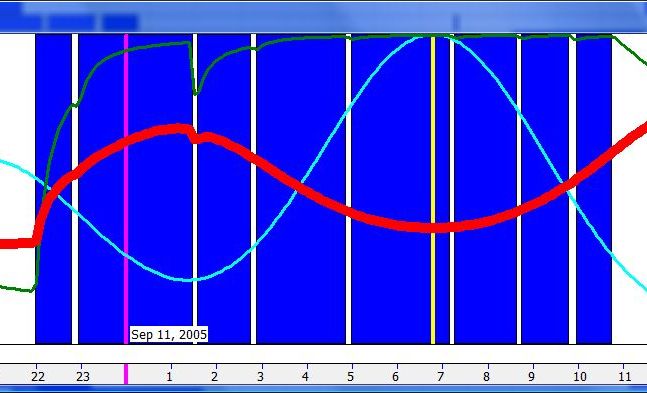
单相睡眠(Monophasic sleep)
单相睡眠图常常显示一个小的午睡时段睡眠倾向峰值,因为即便最纯粹的单相睡眠者也会遇到需要午后小睡缓解的危机日。由于其”危机性质”,这类小睡可能比规律小睡者持续更久。在那些对保护自己自然规律睡眠时段缺乏自律的不规律睡眠者中,中午的峰值尤为明显。
偏好夜间睡眠(Preference for night sleep)
无论先天的昼夜节律周期如何,光对睡眠都有强大影响。尤其是它的相位移动能力,将始终确保人类自然倾向于在夜间睡眠。只有涉及用电和 24 小时工作周期的生活方式的出现,才引发了当前的睡眠障碍流行病,这间接促成了”Uberman 睡眠”等概念的吸引力。
对夜间睡眠的偏好,在不规律睡眠者(尤其是那些患有相位移动障碍并自由运行睡眠的人,或那些在自由睡眠作息下”选择”相位移动(即不试图适应任何特定睡眠时段)的人)中最为明显。在这些情况下,使用 SleepChart 的昼夜节律图,我们可以看到夜间对启动和维持睡眠能力的影响。
在所示的昼夜节律图中,我们可以看到在自由睡眠中对夜间睡眠的明显偏好。该图显示,睡眠启动(蓝线)在晚上 7 点到凌晨 4 点之间更容易,而睡眠长度(红线)在晚上 10 点到凌晨 5 点之间启动睡眠时最大。
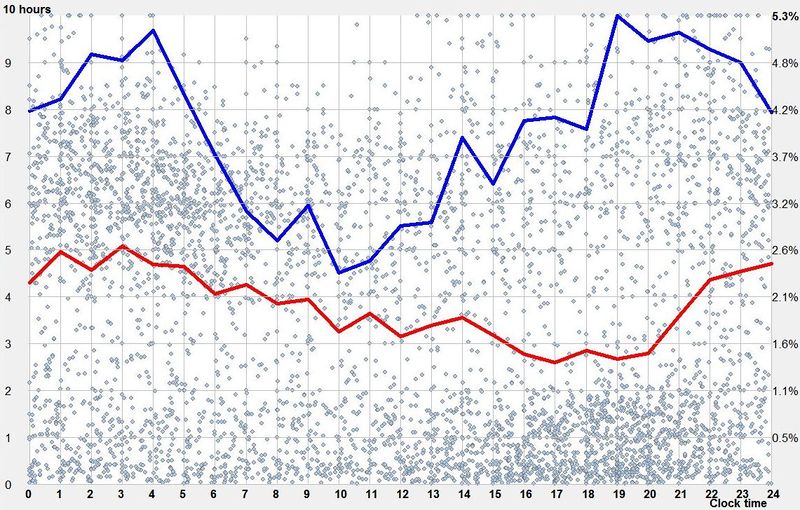
该图也可以被解读为相空间。它展示了在高效的自由睡眠模式中要实现”浪费的”8 小时睡眠有多困难。它也可以用来证明,相空间中没有轨迹会导向驯化的多相睡眠。当闹钟和/或推迟睡眠被引入系统时,睡眠控制可能变得混乱。然而,在自由运行模式下,它会迅速稳定在大致双相的节律周围,通常有一定程度的相位移动,取决于生活方式。相位移动、兴奋性和抑制性刺激的时机,即便它们是重复且规律的,仍可能导致系统出现一定程度的混乱。这发生在刺激周期与驯化的昼夜节律周期不同的情况下。
与第一张图相比,第二个例子可以用来论证,人工照明在具有更规律周期、更好遵守自由睡眠规则的自律睡眠者中,几乎可以消除自然光对周期的影响。
夜间睡眠偏好是否在很大程度上由社会驯化造成,这个问题仍然悬而未决。尽管我们生活在一个 24/7 的社会,白天或晚上的乐趣仍然多于夜间(此时大多数人仍在睡觉)。一个重要线索来自这样一个事实:尽管一天中不同时段的睡眠启动偏好差异不大,但在傍晚或夜间(晚上 8 点 - 早上 6 点)启动的睡眠,仍可能比下午 3 点启动的睡眠长两倍。
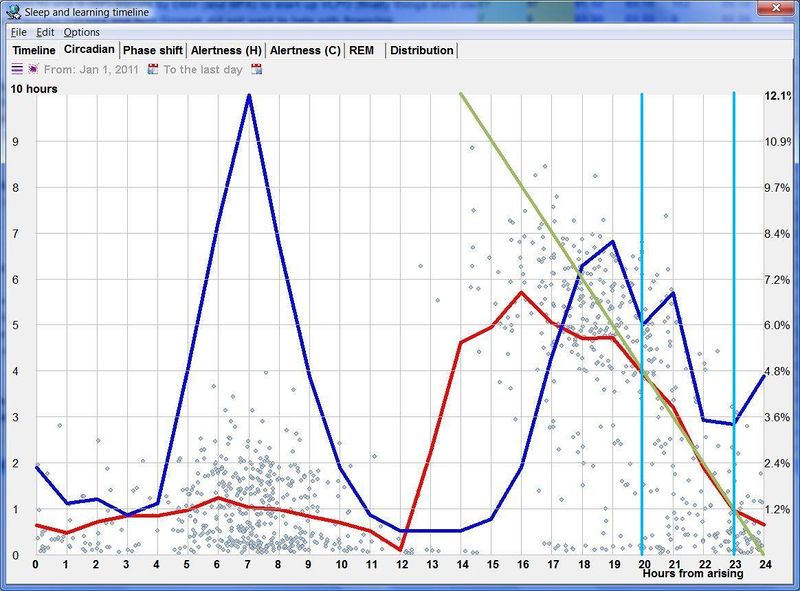
人类睡眠的双相本性(Biphasic nature of human sleep)
大多数研究者同意,人类成年人的昼夜节律周期是双相的(biphasic)。除了睡眠之外,昼夜节律周期的一个外在表现是核心体温的变化。
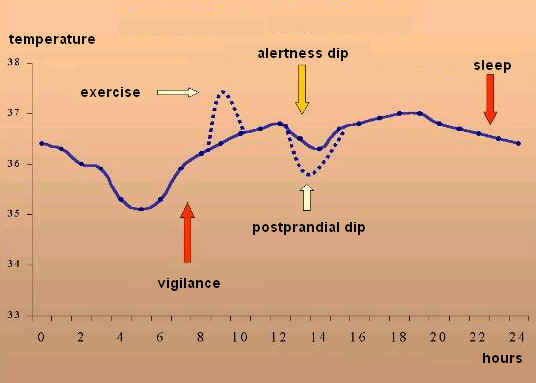
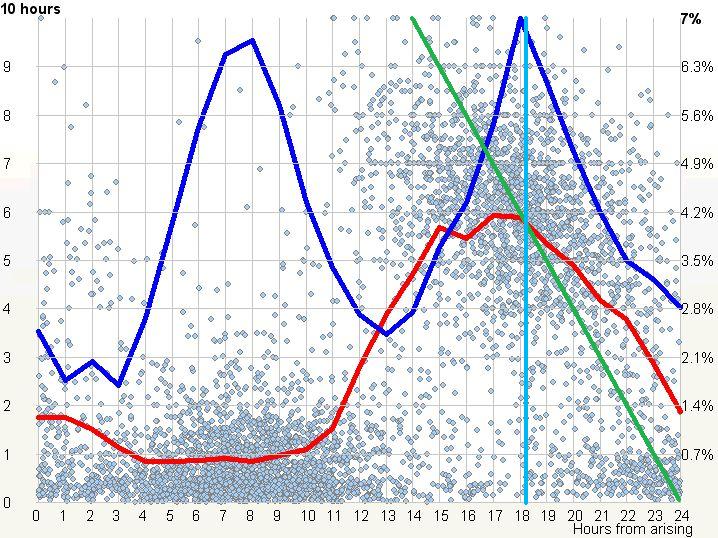
一天中以摄氏度为单位的体温变化(由巴西坎皮纳斯州立大学的 Luiz Menna-Barreto 博士提供)
SuperMemo 和 SleepChart 提供了绝佳的工具来验证人类睡眠-清醒周期双相本性的主张。我收集了来自单相和双相睡眠者的数据,说明了我们的双相本性。
双相学习(Biphasic learning)
仅凭 SuperMemo 就可以通过绘制成绩与时钟时间的关系(无需包含睡眠日志数据)来看到一天中学习表现的双相特性。在本例中,一位单相睡眠者(一个忙碌的两个孩子的父亲)在大约早上 6 点、即自然醒来后不久,显示最佳的学习表现。在上午 11 点到下午 1 点(时钟时间)的学习平均成绩有明显低谷。在下午 5-6 点左右,学习质量有第二次高峰:
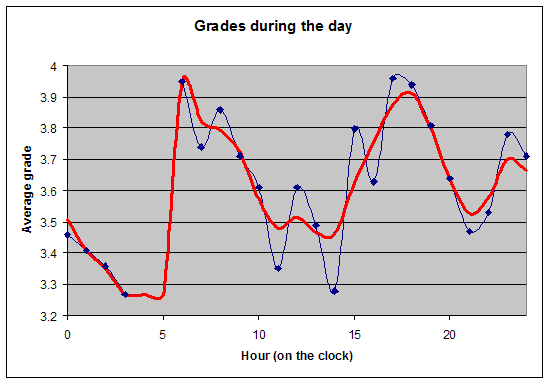
双相睡眠周期图(Biphasic sleep periodogram)
仅凭 SleepChart 也可以用来证明睡眠的双相性。自由睡眠日志可以经过傅里叶分析(Fourier analysis)来揭示睡眠周期性的本质。图中展示了一个示例性的周期图:
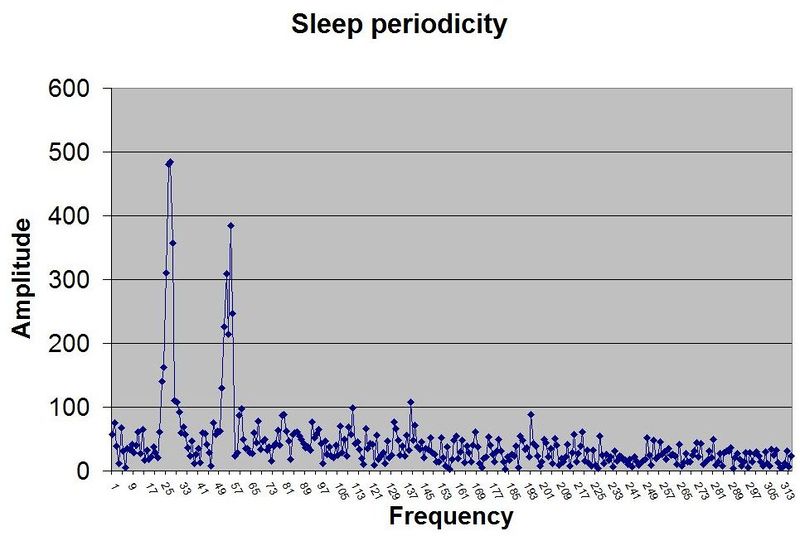
人类自由睡眠的示例周期图揭示了睡眠周期性的双相本性。两个基本睡眠频率主导了这个特定的睡眠日志,它们大致对应 12 和 24 小时的周期。
双相学习与睡眠(Biphasic learning and sleep)
如果我们同时使用 SleepChart 和 SuperMemo,还可以看到清醒表现如何随睡眠相位变化。来自 SuperMemo 的双相成绩图(如前所示)可以根据昼夜节律相位进行修正,而该相位在自由睡眠中可能与实际时钟时间相当独立。在本例中,一位双相睡眠者(睡眠模式不规律的中年男性)在清晨(大约在主观夜晚的估计结束处)显示最佳学习表现:
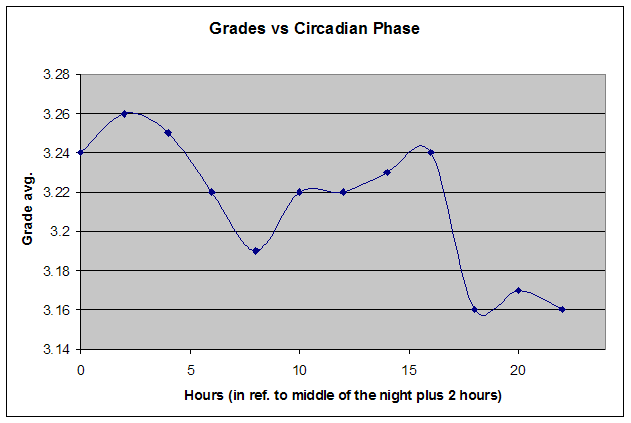
在早晨峰值之后约 7 小时,学习平均成绩有明显低谷。傍晚学习质量有第二次高峰。最后,睡眠前不久学习质量急剧下降。
SuperMemo 中的双相图(Biphasic graphs in SuperMemo)
最新版的 SuperMemo 可以把回忆与由 SleepChart(已与程序集成)估计的昼夜节律相位相关联。在本例中,一位 45 岁的双相男性在一天中显示两个主要的警觉性和学习质量峰值:
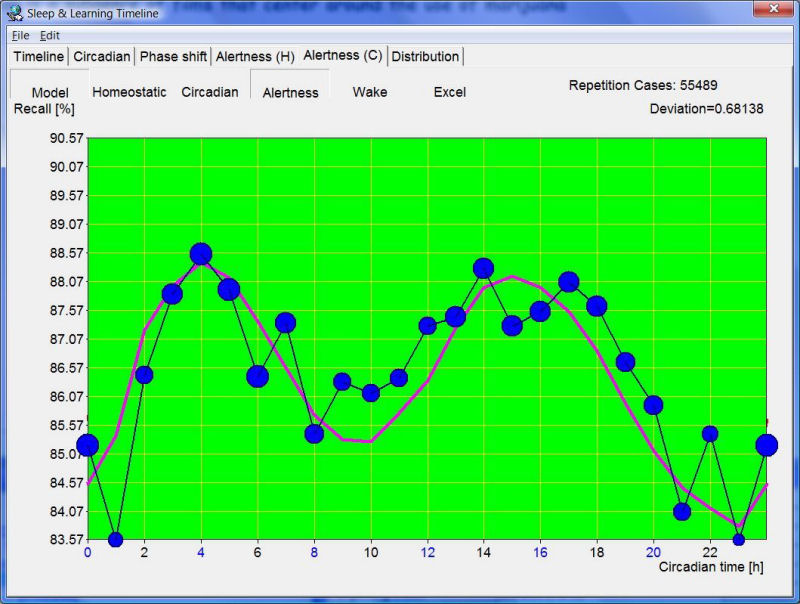
第一个峰值出现在估计自然醒来时间(即实际醒来时间,可能不同)之后的 3-4 小时。第二个、略长的峰值跨越第 12-18 小时。在主观白天时段(即从昼夜节律数据估计的清醒时间,而非实际清醒时间)的第 8 小时,自由回忆有明显的下降。红线显示从 SleepChart 的双组分模型推导的估计总体警觉性。在本例中,估计的警觉性几乎完美地匹配了在实际学习过程中测量的回忆。
带双相学习的单相睡眠(Monophasic sleep with biphasic learning)
对于单相睡眠者,两个警觉性峰值的高度可能不同,他们也会在主观清醒日的第 8 小时左右显示同样的回忆下降。然而,单相睡眠者由于内稳态睡眠驱力组分的影响,可能无法像双相睡眠者那样在傍晚获得同样的表现提升。即便几分钟的小睡也能带来警觉性的重大提升。这一点已被著名的小睡专家 David Dinges 博士在其比较规律小睡者与不小睡者的综合调查中注意到(Dinges,198923)。
为了说明双相和单相睡眠者之间的差异,请看一个类似的回忆图:一位 15 岁的单相不小睡者在早晨时段显示最佳表现,在第 8 个清醒小时有明显低谷,与主观认知功能下降重合:
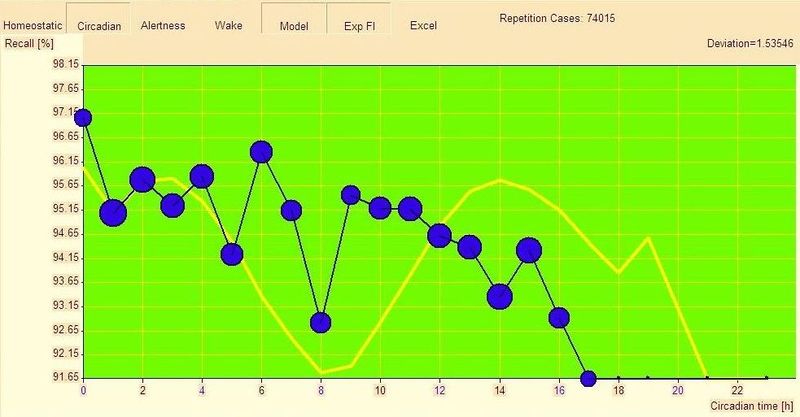
在短暂的低谷之后,出现急剧恢复,然后在下半天表现逐渐下降。这种下降由内稳态机制强烈加速。黄线显示估计的警觉性昼夜节律组分。在本例中,昼夜节律的益处被未在图中显示的内稳态警觉性下降所掩盖。这就是为什么假设的昼夜节律警觉性与实际警觉性只在一天的前半部分匹配。
双相昼夜节律图(Biphasic circadian graph)
睡眠倾向的双过程模型有一个双相的转折。在自由睡眠中(睡眠是睡眠倾向的真实表达),可以在昼夜节律图中可视化睡眠的内稳态和昼夜节律组分:
在规律小睡者中,昼夜节律的双相本性矛盾地由双峰的睡眠倾向曲线而非昼夜节律曲线表达。这种角色反转的原因是睡眠倾向两个昼夜节律峰值之间的生理差异。在规律小睡者中,午睡时段和夜间一样容易启动睡眠。然而,午睡时段的睡眠长度非常短(通常 15-80 分钟)。
在本图中,蓝线对应于在任何给定昼夜节律时间启动睡眠的能力。横轴上,它与 SuperMemo 中显示的警觉性图(如前面段落所示)对齐良好。它与学习数据以及 SleepChart 实现的双过程睡眠模型都对齐良好。
红线对应于维持睡眠的能力。它揭示了之前警觉性图中不可见的内容:午睡不能持续很久,且总是受制于早早的自然终止(第一个蓝色峰值下的低红线)。相比之下,主观夜晚时段是一天中睡眠能够且应该持续最长(通常不少于 4-5 小时)的唯一时段。红色峰值也解释了为什么多相睡眠者渴求”核心睡眠”、醒来时昏沉,并需要重型闹钟武器在这个关键的主观夜晚时段醒来。
David Dinges 在其调查中注意到,一天小睡超过一次极为罕见。大多数小睡者的小睡持续 15-120 分钟。如果在午睡峰值之前小睡,时间会更短;如果在峰值之后,通常更长,甚至可能在特别大的延迟或有睡眠剥夺、REM 睡眠亏欠或任何其他形式的”睡眠债”时与夜间睡眠整合。
Dinges 注意到,无论是食欲性(习惯性)还是替代性(代偿性)小睡者,都倾向于在醒来后 7-8 小时小睡(见:Best timing of naps)。尽管小睡习惯可能不同,但午睡低谷的昼夜节律时机在整个人口中似乎相当相似(Dinges,199224)。
需要再次强调的是,傍晚警觉性的提升被小睡放大,但在不小睡者中也会出现,并可以在双过程模型中轻松解卷积为其内稳态和昼夜节律组分(如接下来两个例子所示)。
双相睡眠倾向的两个组分(Two components of biphasic sleep propensity)
最后两张图显示昼夜节律和内稳态组分对警觉性的影响。
在第一个例子中,一位自由运行的 29 岁不大睡女性在醒来后 8-9 小时显示警觉性低谷。红色内稳态估计没有低谷,在清醒日中稳步下降:
黄色昼夜节律估计显示了低谷的预期位置以及解释傍晚学习表现提升的傍晚峰值:
傍晚回忆提升和傍晚昼夜节律估计对齐良好,再次表明总体警觉性取决于睡眠控制系统的内稳态和昼夜节律组分。
睡眠剥夺中的双相表现(Biphasic performance in sleep deprivation)
中午低谷在严重和轻度睡眠剥夺条件下都同样显著。这张图显示了一项 26 小时睡眠剥夺研究中中午警觉性低谷(Czeisler 等,200625)。低谷的时机(清醒期的第 10-12 小时)表明,前一次睡眠的定位并非最优(因此研究需要打断睡眠)。在实验室条件下,自然醒来可能比强制醒来晚 1-2 小时。该图还显示,由第 2 阶段 NREM 或 REM 睡眠的强制醒来引起的睡眠惰性,比 26 小时的睡眠剥夺造成更大的认知下降。
总结:小睡是好事!(Summary: Napping is good!)
- 人类本性是双相的,并在主观傍晚时段显示学习能力的昼夜节律提升。
- 不大睡者表现出中午表现低谷,也可能从午睡中受益。
分段睡眠(Segmented sleep)
1992 年,Thomas Wehr 博士发表了他在延长黑暗时段(例如北极冬季因纽特人的情况)中睡眠的有趣实验结果(Wehr,199226)。他把实验日分为 10 小时光照(photoperiod)和 14 小时黑暗(scotoperiod)。这种人工控制的环境产生了分段睡眠(segmented sleep),通常由两段 4-5 小时、中间隔着一小时清醒的片段组成。Wehr 发现,睡眠的开始与褪黑素的增加有关,褪黑素在黑暗时段释放(在昼行和夜行动物中皆是)。他还注意到,夜间褪黑素的释放期在较短的照光期中更长(Wehr 等,199327)。
在 10 小时照光期条件下产生的分段睡眠日志片段。
分段睡眠的解释(Interpretation of segmented sleep)
当实验结果被公布时,失眠者欣喜若狂:也许这是正常的?也许我们都应该这样睡? 那些倾向于在夜里醒来、花一小时左右阅读、看电视或扫荡冰箱的人,不再需要感到不正常。确实,区分健康睡眠与不健康模式的最佳标准是睡眠令人神清气爽的力量。只要夜间醒来不导致早晨的痛苦,它们就不应被在意。
历史学家 Roger Ekirch 博士注意到,这种延长的两段式睡眠在电力出现之前的历史记录中频繁出现:“直到近代早期结束,西欧人在大多数夜晚经历两段主要的睡眠,中间隔着一小时或更长的安静清醒。最初的那段睡眠通常被称为’第一段睡眠’(first sleep),或较少见地称为’第一觉’(first nap)或’死睡’(dead sleep)。“(Ekirch,200128)。
睡眠研究者推测,这也许是健康睡眠应有的样子,而我们那些只有单一夜间昼夜节律峰值的睡眠控制模型是错误的。演化论者推测,这可能是对夜间性行为、哺乳或额外警戒时段的适应。Horne 博士喜欢把分段睡眠作为人类倾向于过度睡眠的例子。
分段睡眠与 Borbély 模型(Segmented sleep and Borbely model)
我不同意迄今为止提出的大多数解释,除了那些与主流睡眠研究一致的。我们需要注意,人类和前人类演化的大部分发生在热带地区,那里的夜晚远比北方冬天的夜晚短;从数学上讲,不存在比在每个完整 NREM-REM 周期结束时进入浅睡眠或醒来更偏好”在半夜醒来 1-2 小时”的理由。在 Wehr 实验中观察到的分段睡眠,很容易用睡眠的 Borbély 模型来解释。尽管 Borbély 模型为启动睡眠提供了非常具体的数学条件,但我们必须记住,它只是现实的近似,不一定考虑照明水平或外部唤醒刺激。在延长黑暗和寂静的时段,启动睡眠所需的总体睡眠倾向较低。我将试图用 SleepChart 的双组分模型和一些真实生活例子来阐明我的主张。
分段睡眠与双组分模型(Segmented sleep and two-component model)
当 Wehr 的数据用 SleepChart 的双组分模型处理时,我们看到,睡眠的特征性地在睡眠倾向相对较低的时段启动:
用 SleepChart 采用的睡眠双组分模型解释的 Wehr 分段睡眠。
在现实生活中,双组分模型表明,当警觉性水平降至 33-40% 时启动睡眠。在分段睡眠中,入睡时的警觉性高得多:40-50%。
昼夜节律图显示,偏好的睡眠启动时刻是醒来后第 15 小时,平均产生 5 小时睡眠。由于清醒接近昼夜节律顶点,清醒无法持续很久,因为昼夜节律困倦迅速上升。第二段睡眠随后在第 21 小时出现,平均持续 3 小时。因此,睡眠被分成了 5 小时的前段睡眠和 3 小时的”修正”。
基于 Wehr 数据的昼夜节律图。
睡眠周期图显示在 24 小时周期处有一个典型的频率峰值,以及在 8 和 6 小时处有两个非典型峰值(而非通常的 12 小时午睡峰值)。
基于 Wehr 数据的周期图。
分段睡眠的例子(Examples of segmented sleep)
我在多年的睡眠日志中搜寻自然的分段睡眠例子。我找到的不多。无疑,我可以把这归咎于现代生活方式,它很少允许悠闲的早睡。下面我列出三个来自真实日志的非常不同的例子。几种特征似乎与分段睡眠相关:
- 这种睡眠常常产生高警觉性,例如白天不小睡所证明的。
- 如预测的那样,这种睡眠在冬季更容易找到。
- 强度大的运动可能是诱导向分段睡眠的因素(也许是由于更早入睡)。
- 与 Wehr 的数据一样,SleepChart 的睡眠倾向双组分模型显示,分段睡眠早期睡眠倾向非常快速地下降,这是由于睡眠相对于昼夜节律顶点启动过早。
例子:过早的入睡时间
典型的长黑暗过早入睡的分段睡眠。睡眠启动过早,再次出现非常明显的睡眠倾向下降,导致夜间醒来:
例子:压力导致的夜间醒来
压力导致的过早醒来。带”修正”的分段夜晚:
例子:强度大的运动
强度大的运动诱发、伴随睡眠需求增加的延长睡眠。超过 12 小时的分段睡眠早启动,伴随多次无害的醒来,前 3-4 小时睡眠倾向(红线倒数)快速下降,导致浅而长的睡眠:
分段睡眠的应用(Application of segmented sleep)
这种睡眠产生非常令人神清气爽的夜晚,然而,与现代生活方式相协调地实施它会相当困难。当然,它不会优化时间使用。我只能猜测,把睡眠与昼夜节律顶点良好匹配也应提高睡眠效率。这应该可以用 SuperMemo 数据验证,然而,截至撰写时,我尚未进行必要的计算。考虑到这种古老睡眠对情绪和警觉性的良好效果,我很想让自己接受 14 小时黑暗的实验,然而,10 小时的工作日即便是短期也相当难熬。我也怀疑自己能否压制住一天的想法而早睡。哪怕为实验收集令人兴奋的数据这件事本身,都会让我思绪万千、无法入睡。我也许会在退休后、当我的精力足够衰退、使这成为真正享受时再尝试。
延迟睡眠相位综合征(Delayed Sleep Phase Syndrome, DSPS)
当每天越来越晚入睡的倾向非常强烈时,它可能成为一个严重问题。昼夜节律周期特别长或对授时因子敏感度不足的人,被归类为延迟睡眠相位综合征(Delayed Sleep Phase Syndrome,简称 DSPS)。有时使用缩写 DSPD,其中 syndrome 被 disorder 替代。术语非 24 小时睡眠/清醒综合征(N24、N-24、Non-24)或超昼夜综合征(hypernychthemeral syndrome,有几种拼写变体)偶尔被用来指代最严重的病例。我将始终使用 DSPS 这一标签,以强调这些都是同一问题在不同情况下的不同表达。这与其它作者使用的既定定义相悖,我无礼地不予理会,因为既定术语导致了有害的混淆和一种令人丧失信心的必然感。
在 DSPS 中,个体发现很难在深夜入睡,如果不被叫醒,会一直睡到下午。DSPS 直到 1982 年才被描述,但越来越多的数据表明,各种程度的 DSPS 以流行病的频率发生,尤其是在高中和大学生中。DSPS 个体常常喜欢一直学习到深夜,非常晚入睡(例如凌晨 4-6 点),并发现很难规律地早起。例如,规律地在早上 7 点起床对 DSPS 患者来说是纯粹的折磨。他们常常无法胜任需要清晨表现的工作。他们往往把白天睡眠分成两部分:例如,DSPS 学生常常在夜里睡一小会儿,被闹钟叫醒去上学,在学校里半梦半醒、表现糟糕,放学后美美地睡一觉,直到傍晚才恢复活力和全部脑力。DSPS 学生在午夜之后感觉最好,此时其他人都睡了,他们可以专注于学习或其他活动(阅读、互联网、看电视、电脑游戏等)。
导致 DSPS 的主要因素:
- 生物钟周期增加(远超 25 小时)
- 对重置或前移生物钟的因素(如光、活动、压力、运动等)敏感度降低或增加
- 电照明、24 小时经济及由此产生的”想做更多”的生活方式
正常个体的生物钟周期略长于 24 小时,通过清晨的活动和强光重置。然而,DSPS 个体的时钟周期可能长到难以适应 24 小时。他们还通过傍晚的活动把时钟往前推(与清晨重置同步相反的过程)。DSPS 个体如果被允许在想睡的时候睡,会倾向于越来越晚入睡,也越来越晚醒来。DSPS 患者如果在自己喜欢的时段睡眠,则没有睡眠问题。大多数轻度 DSPS 病例可以通过改变生活方式来改善,但这些改变对患者来说很少是无痛的。如果这一描述符合你的问题,你可以用 SleepChart 免费版诊断你的 DSPS 程度。
青少年中的 DSPS(DSPS in teenagers)
研究表明,DSPS 在青春期非常普遍(Carskadon,199529)。患有 DSPS 的青少年常常发现难以适应正常的上学时间。他们会在上午 10 点(上课中段)体验到最大白天困倦,并在放学后出现警觉性峰值。对许多天然倾向于晚睡的青少年来说,学校成了一种折磨和纯粹浪费时间!教育工作者已经开始关注这个问题;然而,上课时打瞌睡的学生仍是常态!困倦的学生学得很少,并可能自然地对学校整体产生强烈的负面情绪。这是一个规模巨大的问题!如果你是难以起床上学的青少年的家长,你需要现在就行动!否则,这个年轻人的学校岁月将是一段巨大浪费的时间!仅仅要求早睡是不够的。如果你禁止晚间上网,你只是把一剂晚间教育换成了在床上无所事事的辗转反侧。实际上只有一个简单的解决方案:让孩子在自然时间起床,但……在大多数情况下这不现实。你的睡眠治疗师可能也帮不上忙。整个学校系统可能都需要改变,以适应青少年中 DSPS 的普遍性。一些决定把上课时间推迟 1-2 小时的学校已经取得了积极效果。然而,长的昼夜节律周期可能导致学生从长远来看熬夜更晚。研究者建议稳定作息并逐步重新对齐。这些措施仍可能基本上无效。在家上学(Homeschooling)和自由睡眠对这些孩子可能是一个很好的选择。
DSPS 问题的解决方案(Solution to the DSPS problem)
自由睡眠通常是 DSPS 睡眠问题的即时解决方案,然而,它不可避免地会导致”移动”的睡眠模式(见下文)。除自由睡眠外,最著名的疗法是:
- 把睡眠相位循环到与期望时段对齐,以及
- 努力重置周期(见:Curing DSPS and insomnia)。
换言之,如果可能,你可以利用自己每天晚睡 1-2 小时的自然倾向,直到与期望的睡眠节律对齐。此时,真正的战斗开始:努力提供强烈的清晨重置刺激(例如强光、压力、高强度运动等)。这些可以通过傍晚的措施来增强,如褪黑素,以及避免相位延迟因素如光、刺激、压力、互联网等。一般来说,你需要在清晨提供重置刺激,并避免傍晚的延迟因素如电脑、电视、人工照明等。对大多数人来说,一定程度的睡眠剥夺比傍晚在黑屋里无所事事几个小时更容易接受。
DSPS 是一种疾病吗?(Is DSPS a disease?)
也许,大多数 DSPS 病例可以用基因决定的睡眠控制系统与生活方式之间的不兼容来解释。对某些人,问题程度比其他人更大(见:生物钟基因及影响时钟周期的突变(Golombek 和 Rosenstein,201030))。每个人都可以通过决定彻底改变习惯(例如回到农民的生活方式)来轻松治愈这一障碍。然而,由于就业类型或家庭生活条件,这种改变通常不可行。这意味着 DSPS 患者在一段时间内,注定要与其生物钟进行持续战斗。
官方数据显示 0.2% 的成年人患有 DSPS。利用大量 SleepChart 数据提交,我看到了 DSPS 的真正流行。此外,还有一个庞大的隐性 DSPS 人群。我见过这样的案例:一些人在闹钟下、看似正常但略受睡眠剥夺污染的正常作息下生活多年,一旦放弃闹钟就开始显示出 DSPS 睡眠模式。
诚然,给我写信的人已经是预选人群,但数字确实惊人。我相当确信,这些 DSPS 病例大多与生活方式有关。由于”综合征”一词可能暗示 DSPS 是一种疾病,我一直强调,DSPS 更是我们现代以电为基础的生活方式的反映,而非一种真正的障碍。有趣的是,我收到的 ASPS 提交很少。似乎是 DSPS 患者才会熬夜在电脑上搜索睡眠问题的解决方案,最终到达 SleepChart 和自由睡眠的概念——这可以是他们的神奇解药(如果真能负担得起的话)。
DSPS 流行病可以被视为一种文明障碍,现代生活方式的压力与数百万年的演化相悖。从长远来看,一旦我们完全理解睡眠背后的所有生化和激素过程,温和的药物干预可能使调节昼夜节律周期成为可能。
异步 DSPS(Asynchronous DSPS)
对抗相位移动(Combating phase shift)
患有 DSPS 的人发现难以与 24 小时时钟同步。在下面的图中,一位患有轻度 DSPS 的青少年女性,由于与”世界其他部分”同步的失败努力,睡眠节律发生了解体:
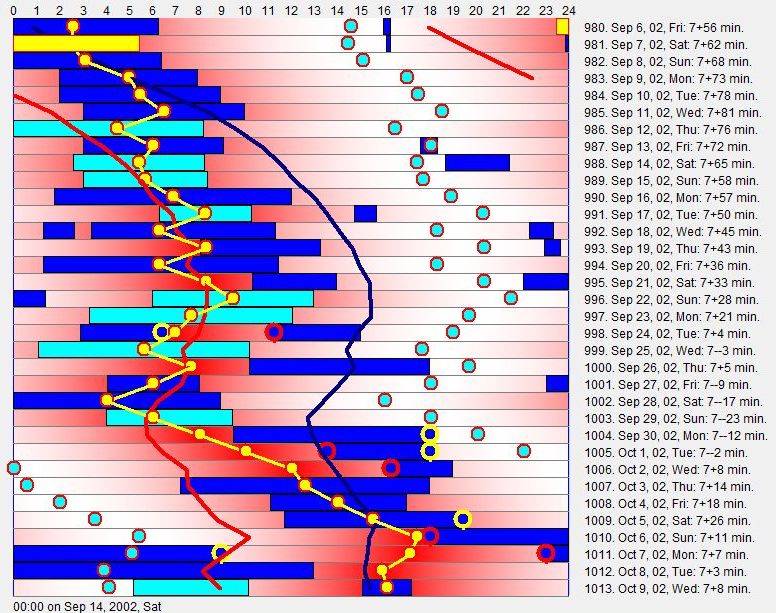
假期结束后,她在 9 月初开始时与世界其他部分同步良好,在 6:30 到 10:00 之间醒来。然而,她的生物钟在主观夜晚时段持续移动。很快她在 12:00 醒来,并开始对抗进一步的移动。这导致睡眠周期的解体、低于偏好平均(例如 4 小时)的短睡眠时段和频繁的疲倦发作。SleepChart 试图绘制主观夜晚(即最大自然困倦时段)的范围。统计预测的主观夜晚被红线和蓝线括起。昼夜节律顶点(夜间中点)以黄色绘制。昼夜节律睡眠倾向以红色深浅表示。被闹钟终止的睡眠区块以青色标记。显然,睡眠模式的最大破坏发生在”自然”节律与”期望”节律偏离最远的点。轻度 DSPS 病例能够以与困倦持续斗争为代价,迫使生物钟大致保持在期望时段。在更严重的病例中,昼夜节律变量将运行 24 小时周期,当自由运行变量再次与”期望”睡眠时段对齐时,个体会体验到”好睡眠”的回归。
平均睡眠长度为 6.8 小时,但总睡眠日复一日变化很大。由于与自然节律的斗争,平均 DSPS 移动难以确定。然而,移动可能在 60 分钟左右,这由昼夜节律顶点估计(黄色)的平均进展所证明。如果不使用闹钟,前进的睡眠相位可能在 3-4 周内完成完整的 24 小时回转。
重新同步周期(Resynchronizing the cycle)
在下一个例子中,一位患有严重 DSPS 的中年女性,在将睡眠节律稳定在社会可接受范围内时经历了类似的斗争:
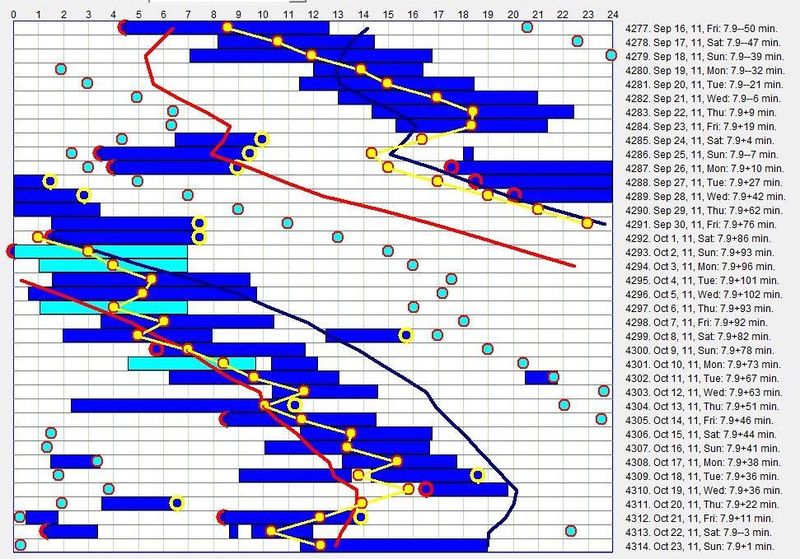
当身体屈从于睡眠相位的进展(9 月 16 日 - 9 月 23 日)时,主观困倦最小;在与进展的斗争(10 月 2 日 - 10 月 11 日)期间,白天疲倦明显增加,其中浅青蓝色的睡眠区块是被闹钟人为终止的区块。最后,在睡眠时段与昼夜节律相位失去同步(10 月 19 日 - 10 月 22 日)期间,白天困倦达到峰值。突破来自对自由睡眠的一丝不苟遵守。下一份日志显示同一女性在管理良好的自由睡眠作息下:
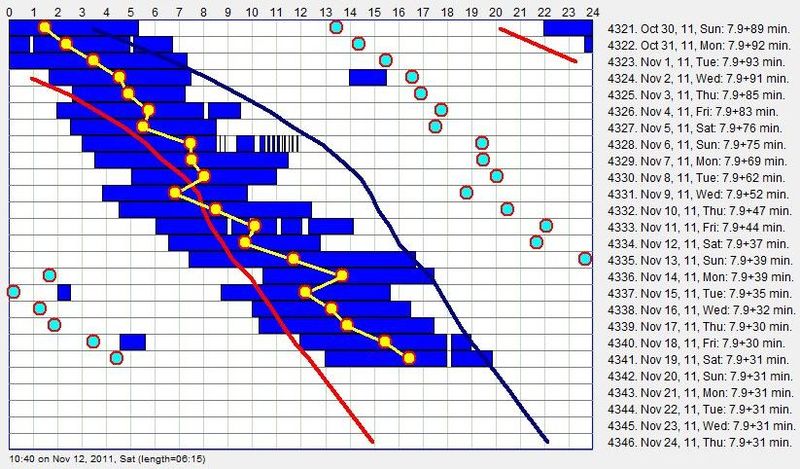
睡眠时段与昼夜节律顶点(主观夜晚中部的黄线)的完美对齐,带来了三倍的能量和幸福感。
DSPS 中的社交生活(Social life in DSPS)
自然地,只有远程办公、自雇或在家办公的人才能负担在 DSPS 中让睡眠自由运行。即便如此,移动的睡眠相位也是一个严重的困境。一位来自荷兰的法定盲人 DSPS 患者写到移动睡眠模式的痛苦:“我在自由睡眠。我有一个 17:30 的预约。我预计像前三天一样在 15:00 左右醒来。结果我在 17:00 左右醒来,还有点累。我不得不跳过早晨的例行公事(冥想、早餐、SuperMemo 等)。自由睡眠对我非常有效。但今天糟透了。因为比预期晚醒而不得不匆忙赶路,真的很紧张。” 经过医疗咨询,这位受试者被开了晚间褪黑素,能够稳定周期(至少在撰写本文时持续了几周)。同一受试者在自由运行和使用褪黑素之前与相位移动的痛苦斗争,见此图:
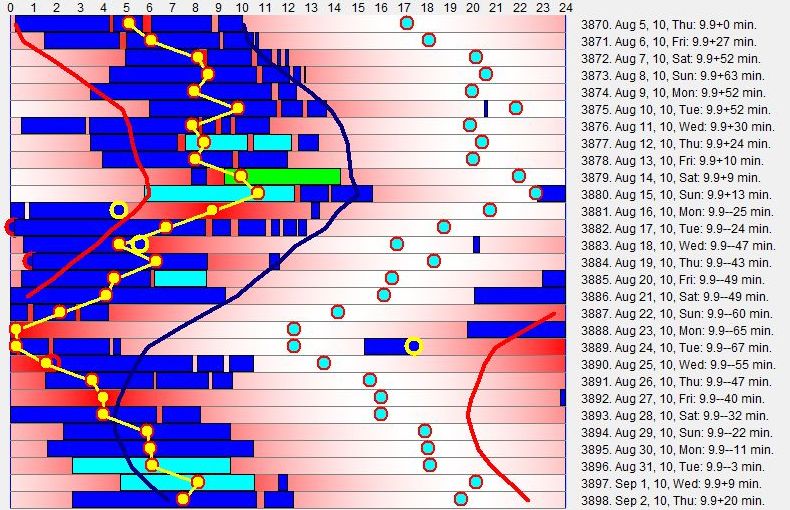
这个例子说明了所有更严重 DSPS 病例的主要困境。自由睡眠常常会产生相位移动。任何倾向于很晚醒来的人,也很可能在自由睡眠中每天越来越晚醒来。这是 DSPS 的标志症状。然而严重的 DSPS,只要睡眠自由运行,就其本身而言从来不是健康问题。最令人烦恼的是日程安排问题。选择介于两个极端之间:
- 要么让自己的生活更少依赖于可能与睡眠作息冲突的会议和约会,或
- 研究 DSPS 疗法以稳定睡眠-清醒周期。
如果你碰巧总是很晚醒来,那么总是在同一时间醒来会让日程安排容易得多。如果你不选择上述两个极端之一(自由作息 vs. 稳定战斗),你将冒着使生活相当艰难的冲突风险。更危险的是,如果在自由睡眠作息下打乱昼夜节律,各昼夜节律变量之间可能失去同步。这将导致一种情况:在一天甚至几天内,你不确定最佳入睡时间。即便 SleepChart 也可能无法做出良好预测。这不可避免地导致睡眠质量差和几天低生产力。
DSPS:基因还是生活方式?(DSPS: genes or lifestyle?)
尽管我反复说 DSPS 流行病是现代生活方式的反映,但基因因素显然发挥作用,“创造性人格”也可能是问题的根源。这里有一个有趣的故事:一位作家妈妈,她的父母和两个孩子没有任何睡眠问题的迹象,然而她患有严重的 DSPS,她 20 岁的儿子也是如此:
“我母亲声称我从出生那天起就有睡眠问题。在那些日子里’母婴同室’是新鲜事,因为大多数婴儿都被放在婴儿室里,正常分娩后母婴在医院住一周是常态。她津津乐道地讲述护士如何迫使她让我’母婴同室’,因为’我整夜让婴儿室里所有其他婴儿都睡不着’,因为我拒绝在夜间时段入睡。她说她和我父亲把我带回家后,我在他们房间睡了一晚,但我整夜都醒着,所以从那以后我不得不’哭到睡着’,因为我就是不在夜里睡。当我几岁时,有一天晚上,我父母无奈地说:‘好吧。如果你想熬夜,你可以坐在这里看 Johnny Carson 节目。‘显然我很乐意这样做!然而,我有非常不愉快的记忆,可以追溯到我有记忆之初:每天晚上大约 20:00 被放上床,在黑暗无聊的房间里,清醒地、无所事事地躺着好几个小时。那时候没有互联网、没有有线电视、没有电子游戏、没有手机等等。我一生中从来都无法在体面的时间入睡!我常常唱歌哄自己入睡,这要花好几个小时。有时我唱完所有的歌,不得不重复几首直到昏睡过去。”
如今,这位自雇女性正在实验自由睡眠,并声称,除了因此受损的社交生活外,新作息把她从”持续困倦的地狱”中拉了回来。
时间疗法及其风险(Chronotherapy and its perils)
时间疗法使把昼夜节律相位塞进期望时段变得容易,例如在洲际飞行后、在昼夜节律障碍中,或为了轮班工作。对大多数人来说,通过诱导相位延迟向前移动周期更容易。每天晚睡 40-70 分钟,循环穿过白天,直到达到期望的睡眠相位,是可能的。推动昼夜节律周期应始终是最后的手段。如果可能,所有人为的睡眠控制形式都应避免,因为它们对健康并非中性。然而,文献中的一些报告表明,时间疗法可能有严重的长期周期同步后果。Wehr 报告(Wehr 等,199226):“1983 年,我们中的一位描述了一位 28 岁、患有 DSPS 的男性,他接受了时间疗法后,发现自己的睡眠时段无法停止绕时钟旋转,也无法把节律恢复到 24 小时作息。相反,发展出了超昼夜综合征,持续的 25 小时睡眠-清醒周期。这种罕见综合征极其令人衰弱,因为它与大多数社交和职业义务不兼容。”
我亲自目睹了许多看似正常的人在转向自由睡眠时的相位延迟,并有不同的解读。在自由睡眠中维持平衡的 24 小时周期,个体之间困难程度不同。这种困难在昼夜节律图中很好地由睡眠维持曲线相对于决定平衡 24 小时周期的盈亏平衡线的角度表达。如果允许青少年在深夜沉浸于他们的爱好和热情,大多数人都会在一定程度上难以维持平衡。一旦被允许随心所欲,他们常常仅仅因为不愿”按时”上床而诱发相位延迟。即便渐进的 24.5 小时周期一开始可能令人不安、导致在不寻常的时间睡觉,自由运行带来的延长清醒日的便利可能超过负面副作用。对 DSPS 患者,自由运行可能提供一种异常程度的舒适,难以放弃。难怪许多享受自由睡眠中渐进移动昼夜节律周期的 DSPS 患者,常常放弃重置周期的战斗,甚至发现自己的自然相位移动远比最初诊断的大。
由于严重 DSPS 病例的医学和精神病学术语非常混乱,我需要再次尝试解释”不规则的非 24 小时睡眠-清醒综合征”病例,我指的是睡眠事件不落入规律模式的严重 DSPS。我在 Asynchronous DSPS 一节中展示了某些案例。我毫不怀疑,这类不规则大多来自受试者对自己睡眠偏好的无知,以及在昼夜节律障碍中健康自由睡眠作息规则的无知。一旦向患者讲授健康自由睡眠的规则(也许在 SleepChart 的帮助下),睡眠模式就会变得规律。
因此,我个人对时间疗法的立场如下:时间疗法是重新定位昼夜节律周期相位的最佳方法。它应始终是最后手段,因为它对睡眠质量并非中性。然而,偶尔使用温和时间疗法的长期后果可能可以忽略。
同步 DSPS(Synchronous DSPS)
当 DSPS 患者尝试自由睡眠时,睡眠相位延迟是不可避免的。文献中有记录人们沿着这种移动 DSPS 作息生活数十年而无重大健康副作用的案例(Neubauer,200031)。一个示例报告见这个博客32。一些作者声称,移动的作息可能增加抑郁症、酒精中毒或镇静剂依赖的发病率(这是试图在”适当”时间诱导睡眠的结果)。
下图展示了一位自由运行的中年自雇男性的睡眠模式:
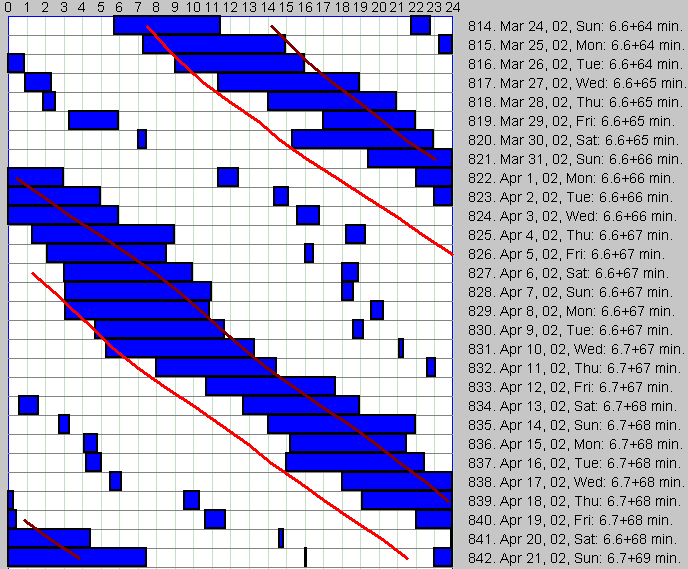
图中可见一个非常清晰、规律的 DSPS 模式,每日相位移动 64-68 分钟。尽管在”不自然”的时段睡眠在健康上肯定不如正常睡眠,但对 DSPS 受试者来说,自由睡眠节律可能远比任何试图适应”标准”生活方式的努力更少压力和破坏。同步 DSPS 的一个非常可靠的判定标志,是睡眠启动时间与睡眠持续时间之间联系的丧失(见:Preference for night sleep)。由于睡眠持续时间由昼夜节律相位决定,良好同步的睡眠作息显示睡眠长度的变化很小(本图中为 6.6 小时)。尤其是,睡眠长度独立于睡眠启动时间。每当受试者试图与日光或与日光相关的活动同步时,睡眠长度与启动时间之间的联系就会重建。DSPS 患者常因晚睡倾向被误称为”猫头鹰”,而 ASPS 患者被称为”百灵鸟”。该图说明了为什么这是一个误称。
类似的图显示了一个相位移动程度更大(84-90 分钟)的 DSPS 案例:
由于在与昼夜节律周期良好对齐的睡眠事件中睡眠效率更高,规律自由 DSPS 作息的人报告的主观警觉性和能量,远高于不规律 DSPS 作息的人。这种差异也出现在 SuperMemo 收集的数据中。
28 小时日作息(28 hour day schedule)
28 小时日作息是为那些寻求更高生产力和一天中更多小时的人提出的。一个 28 小时睡眠模式设计的例子显示,任何人要保持该作息更长时间,每日相位移动需达到 4 小时的极端:

28 小时日作息设计。
28 小时作息的优势据说包括更长的工作日、规律的 6 天周、可重复性、长周末、增加的能量、无限的睡眠等(详见:28 hour day)。这一命题是多相睡眠光谱另一端的命题。然而,它似乎远比多相睡眠容易维持,因为它不需要闹钟。睡眠研究者认为,这一作息如此极端,没有人能在长期维持。4 小时的相位移动如此不可能,以至于研究者选择它用于实验性的强制失步协议。这些是身体本应无法适应授时因子时机的实验协议。这种驯化失败对研究自由运行的昼夜节律变量有益。4 小时移动已被用于前移和延迟协议(20 小时日(Wyatt 等,199933)和 28 小时日(Carskadon 等,199934))。迄今所有研究似乎表明,在强制失步协议中,昼夜节律周期由于重置刺激互相抵消而继续在后台自由运行,睡眠事件以不可预测的模式与昼夜节律变量相交,导致分段睡眠、过早醒来、睡眠缩短、REM 减少和其他异步症状。简而言之,28 小时日被认为极端到足以造成睡眠时机与昼夜节律周期之间永久的失步。
DSPS 中的 28 小时日(28 hour day in DSPS)
一些 DSPS 患者报告在 28 小时作息上比在常规 24 小时作息上感觉更好。我不认为存在天生的约 28 小时昼夜节律周期的个体,然而,可以想象,把 DSPS 周期塞进 24 小时的努力比以把周期拉伸到 28 小时的替代方案更痛苦。主要区别在于,缩短周期通常涉及痛苦地使用闹钟,而拉伸周期”只”需额外的 2-3 小时行尸走肉般的清醒。即便在严重 DSPS 中,要适应 28 小时作息也相当困难,因为相位响应曲线表明,睡眠相位对强烈延迟的入睡时间响应不强,在极端情况下甚至可能导致相位前移。超过 2 小时的相位延迟应极为罕见。
对大多数人来说,即便对自己最佳周期时段的轻微偏离也很难容忍。因此,除非来自对自己睡眠偏好的严格分析,否则所有设计师作息都应避免。同样,自由睡眠是更好的选择,即便它在规划社交或职业生活方面可能更不可预测、更不方便。
让我们考虑受试者 S 的示例案例,并比较她在常规、28 小时和自由睡眠作息上的睡眠效率。
严重 DSPS 中的常规作息(Conventional schedule in severe DSPS)
当 S 试图在医疗监督下、借助睡眠药物(包括褪黑素)坚持常规睡眠作息时,睡眠严重碎片化、短、不令人神清气爽,且作息不可持续:
受试者 S 在不可持续的常规作息上。
这种睡眠无异于精神折磨,所有有类似驯化失败程度的个体,都应始终被允许以严重残障为由让其睡眠自由运行。
在所给的图中,可以在主观夜晚于 2011 年 4 月 6 日左右离开常规夜间时段时,注意到可能的自由运行昼夜节律周期模式。这在睡眠事件合并后更为明显:
受试者 S 在不可持续的常规作息上(睡眠事件合并)。
毋庸多言,常规作息如果维持更久,可能因持续的睡眠剥夺加上药物而导致严重的健康问题。尽管所给案例相当极端,但遭受类似睡眠问题的人数不断增加,肯定影响着整体人口健康和生产力。
严重 DSPS 中的 28 小时作息(28 hour schedule in severe DSPS)
当 S 试图坚持 28 小时日作息时,她的主观睡眠质量连同总睡眠量急剧提高。然而,对睡眠图的详细分析显示,睡眠碎片化仍然相当严重,显示出睡眠控制系统的紧张:
受试者 S 在 28 小时日作息上。
在两个周期后开始出现分段睡眠,这可能表明实际相位移动落后于计划的相位延迟。分段睡眠常是过早入睡时间的标志,出现在 28 小时作息入睡时间早于假定的自由运行主观夜晚时。当睡眠作息最终崩溃时,漫长恢复睡眠事件的定位似乎表明,平均每日相位移动实际上可能远少于 4 小时。在极端情况下,主观夜晚与计划夜晚之间的巨大差异可能导致自我抵消的相位移动,这反而可能稳定睡眠周期。
同一 28 小时日作息的昼夜节律图说明了睡眠控制系统中的混乱程度:
受试者 S 在 28 小时日作息上的昼夜节律图。
严重 DSPS 中的自由睡眠(Free running sleep in severe DSPS)
当审视 S 在仅仅几个月前自由运行的睡眠图时,睡眠相位移动更接近每天仅 41 分钟(而非维持 28 小时日作息所需的 240 分钟):

受试者 S 在自由睡眠作息上。
然而,该图显示,即便在那段时期,睡眠也严重碎片化和不规律。类似分析常常因复合情况(如疾病、家庭问题、药物,甚至错误选择入睡时间(例如试图稳定或加速周期))而具有误导性。
在类似案例中,绘制自己精确的昼夜节律偏好至关重要。因此,几周不间断的自由睡眠对确定自己天然内在的每日相位移动来说非常宝贵。一旦做到这一点,就可以对给定个体 28 小时日作息的可持续性有更多说法。然而,对于那些绝对无法平衡周期、且能负担难以与外部世界节律调和的睡眠作息的人来说,持续的自由睡眠始终是最佳选择。见:Curing DSPS and insomnia。
治愈 DSPS 和失眠(Curing DSPS and insomnia)
如果你不靠闹钟就无法及时醒来上学或上班,你可能患有延迟睡眠相位综合征(DSPS)。DSPS 也与试图保持更早入睡时间时的入睡困难有关。换言之,任何 DSPS 的治愈方法也很可能解决入睡性失眠问题。如果你带着 DSPS 问题去看睡眠专家,你很可能会被开褪黑素或强光疗法,结果却发现它们对睡眠质量的影响有限。如果你是失眠症患者,可能还会被开安眠药,这或许能帮你入睡,却达不到期望的效果:神清气爽的头脑。本章应帮助你解决问题。利用人类睡眠控制系统的特性,可以从数学上证明 DSPS 及相关失眠的问题总是可解的,然而,解法不一定意味着最敏锐的头脑或最高的智力生产力。此外,许多人仍会因缺乏自律而失败!在现代世界入侵人类生物学之处,仍可以用理性的规则抵御潮流。然而,这意味着对健康生活十诫的一丝不苟遵守。生活表明,人类觉得所有十诫都难以遵守!
失眠、DSPS 和 N-24 的解法(Solution for insomnia, DSPS and N-24)
这个算法应在以下所有情况帮助你:
- 如果你是失眠症患者,你的身体很可能试图在不同的相位睡眠,如果你修复相位,你将修复失眠。
- 如果你是 DSPS 病例,尤其是长期患者,你会对这个算法非常怀疑。你需要给它至少几周自律的尝试,才能看到你的睡眠周期可以用正确的工具修改。
- 如果你是 Non-24 且认为你的睡眠没有模式,你很可能是错的。确实存在可能彻底搞乱你睡眠的罕见突变,但更可能的解释是,你是一个 DSPS 病例,只是向睡眠模式中引入了足够的混乱使其看似不可预测。如果你在其他方面健康并尝试自由睡眠,你会注意到模式。要让模式出现,你的”自由”睡眠必须真正自由。没有调节,没有人为控制!之后,你可以尝试下面给出的算法,看看自己是否能展现足够的自律来稳定周期。
DSPS 与生活方式(DSPS and lifestyle)
尽管 DSPS 有各种基因影响,但对大多数人来说,问题很大程度上是生活方式的事。我之所以这样说,是因为回到农民的生活方式可以保证 DSPS 问题的消失。下面,我汇编了一个简单的算法,在足够自律的情况下应能在绝大多数案例中解决 DSPS。到目前为止,在向人们展示如何用自由睡眠应对 DSPS 方面,我比下面列出的处方更成功——后者是自由睡眠的衍生,带有一些针对防止相位延迟的限制。所给出的算法之所以失败,主要是因为一个问题:违反规则!确实存在一些带有精神病学色彩或其他健康问题的真正硬核 DSPS 病例,可能特别棘手,然而,这些应是在日益增长、与 DSPS 挣扎的人群中罕见的少数。这群人现在包括无数的失眠症患者,他们从未听说过 DSPS,由于使用闹钟甚至从未到达相位移动的问题。Weitzman 假设,相当数量的入睡性失眠患者可能患有未诊断的 DSPS(Weitzman 等,198135)。现在我们知道这一假设肯定成立,这可以通过让失眠者自由睡眠来证明。在解除对睡眠时机的限制后的头几天内,可以观察到显著的相位延迟。同时,伴随入睡性失眠几乎瞬间消失。
不可信的解决方案(Implausible solutions)
患有 DSPS 的人常诉诸自己不可信的解决方案,包括:
- (管理不善的)自由睡眠,伴随不可避免的相位延迟,最终导致白天睡觉。
- 在入睡时间达到离谱晚的程度(通常在黎明之后)时跳过一夜睡眠。
- 用药物(包括褪黑素)和人为授时因子(如闹钟或一位把孩子从床上拽起来上学的妈妈)稳定睡眠模式。
- 在不可预测时段发生的混乱睡眠,失去对夜间的感觉,伴随长时间的困倦、低能量或健康不佳。这种睡眠模式常被标记为 Non-24 或 N-24,然而在大多数情况下,它与 DSPS 的区别仅在于缺乏遵守自然睡眠周期所需的自律或知识(即便其周期长于 24 小时)。
只有管理良好的自由睡眠才能在 DSPS 中以最低的负面健康结果风险产生健康的睡眠。然而,很少有实践者真正遵守自己生物钟的规则,因为总有借口或不可避免的理由在主观夜晚与白天义务或消遣冲突时违反它。在 DSPS 中尝试”自由”睡眠多年的人,有搞乱睡眠控制系统的风险。我从他们的睡眠模式常常变得越来越不规律32、睡眠质量常常下降这一事实得出结论。如果睡眠真正自由,没有药物、光疗、人为延迟或使用闹钟,这种效应几乎肯定会被最小化。他们甚至可能错误地声称周期开始越来越长,而实际上是生活方式要求不断拉伸清醒时间。这里的罪魁祸首自然不是自由睡眠本身,而是由于睡眠与白天活动冲突而不可避免的违反。
可信的解决方案(Plausible solutions)
如果我们排除健康的农民生活方式和放弃晚间用电,那么 DSPS 问题只有两种合理的生活方式解决方案:
- (管理良好的)自由睡眠,伴随相位延迟、高创造力、高生产力、良好健康,以及几乎无法与外部世界调和的作息。
- 稳定的 24 小时睡眠周期,伴随僧侣般的自律、良好健康和较低的生产力。
我在全文中都写到自由睡眠。我如此推崇自由睡眠,以至于被指责给 DSPS 盖上了”不治之症”的印记。这里,我想提出一个可信的、在 DSPS 中以最小人为干预维持 24 小时睡眠模式的算法。
算法(The algorithm)
失眠和 DSPS 问题唯一合理的 24 小时解决方案,是改变睡眠相位。我们可以用傍晚措施(把睡眠向后拉)和清晨措施(把睡眠向后推)来前移睡眠相位。这解决了睡眠的昼夜节律组分。此外,所有在傍晚增强内稳态困倦的措施也是受欢迎的。然而,没有昼夜节律组分,这些实际上可能加剧失眠。这就是为什么只有下面给出的综合方法才提供离开 DSPS 和/或失眠的可靠机会:
-
确定一天的长度:用 SleepChart 确定你最佳清醒日长度。你可能需要几周的自由睡眠数据才能做出良好估计。如果你无法自由运行,和/或程序似乎太复杂,你可以做一个有根据的猜测。例如,如果你认为每晚需要 8 小时,在自由睡眠中可能接近 7 小时,最佳清醒时间将是 24-7=17 小时。然而,由于可能的失眠问题,从稍长的一天和较短的睡眠开始是有意义的,例如从 17.5 小时清醒开始。
-
确定偏好的醒来时间:这应是每天同一时间!如果你每周只需早起一次,你的整个周期应被定位以适应那个早间时刻,否则你将面临昼夜节律系统中永无止境的涟漪和可能的失败。如果醒来时刻足够晚以确保清晨阳光,你的机会将大大增加。
-
虔诚的入睡时间:把你的最佳清醒时间加上所选醒来时间,你将得到必须被虔诚尊重的最佳入睡时间。这种尊重是成功的关键之一。过程中你会遇到许多障碍。最令人困惑的之一是,最佳清醒时间会根据一天的内容、活动、压力、天气、季节、社交互动等而变化。对大多数人来说,这些变化少于一小时。然而,你更可能属于剧烈变化类别,因为这是 DSPS 的潜在问题之一。开始把你的最佳入睡时间当作生命攸关来遵守。如果你非常兴奋或紧张,在入睡时间不困,可以在 20-30 分钟后再试。你需要确保不会辗转反侧,也不会过早醒来。如果你看到任何失眠迹象,你就会知道你的入睡时间来得太早,你分配了太多睡眠时间。如果你在入睡时间之前非常困倦,并毫无疑问会睡得像根木头,就早点睡。你的最佳入睡时间是指导,你实际的困倦是最终的衡量标准。
-
启动算法。理想情况下,你应在醒来时间早于期望醒来时间时启动算法。否则,你可能需要暂时诉诸闹钟。然而,每次使用闹钟都会增加你与早睡相关的压力和焦虑。如果你必须使用闹钟,并在早上起床困难,你应该考虑完整地自由运行一个周期,每日延迟 20-40 分钟,直到你能舒适地在所选醒来时间前 2-3 小时起床。然后你可以尝试重新启动算法。
-
保护区:由于你患有 DSPS,你的主要目标是防止相位延迟。这意味着在你最后 2-4 个清醒小时里,你需要避免光、电、兴奋、压力、剧烈运动、电视、电脑、社交互动、电子邮件、网上冲浪等。如果给出的算法长期对你不起作用,你可能不得不把这个”保护区”延长到产生的”不作为”带来的压力或挫败感超过它实际消除的程度。把保护区延长到超过 2-3 小时的黑暗沉默可能是适得其反的。自己看看。记住,保护区不是躺在床上的时候!床上无效的时间会增加你的压力,如果你早睡,你可能搞乱那晚的睡眠并实际上延迟相位!显然,在保护区你能做的活动非常有限。我把它留给你的想象力。也许是昏暗光线下的适度运动?或者性爱?或者散步和思考?Seth Roberts 信奉睡前单腿站立。
-
清晨光照:最强大的授时因子之一是光。如果阳光能在你应该醒来时照进你的房间,你可能消除实现 24 小时平衡周期一半的困难。如果由于季节、早间时刻或居住安排而无法实现,你可以考虑使用”强光疗法”中的商业设备之一,尤其是如果你用定时器开灯(如果你的设备没有定时器,你可以使用一个在夜间切断电源的定时器)。请务必阅读所有相关安全说明,以防止对健康的任何长期影响。
-
清晨运动:另一个强大的授时因子是户外运动。如果可能,你应在清晨或至少在一天的前 5 小时安排运动。清晨运动会影响你的智力表现,你很可能会更快疲倦。然而,这也是运动有助于你睡得更好的原因之一。一天晚些时候的剧烈运动可能有相反的效果,可能延迟你的相位。然而,某些形式的运动不会有这种效果,因为相位移动刺激会被内稳态睡眠驱力的放大和有助于早睡的”禁食效应”所抵消。这些傍晚运动应无压力、无伤害、无情绪。温和的体操、瑜伽或拉伸是无害傍晚运动的例子。如果不过于兴奋或费力,甚至健美也可以。这种晚运动最好在昏暗光线和无其他唤醒刺激的情况下进行。如果你能发誓傍晚运动(即便是剧烈的)帮助你更快入睡,记住睡眠本身有相位移动的力量,实际上可能超过肾上腺素或运动活动的相反效应。如果你不确定,自己实验。
-
傍晚禁食:除非你是那种不吃零食就无法入睡的贪婪型,否则在一天最后几小时放弃用餐。如 DMH 一节所解释的,食物可能对你的昼夜节律周期定位有额外影响,你不想让你的大脑认为傍晚是获取食物的适当时机。早晨健康的早餐应对睡眠相位产生相反的效果,并总是被营养学家推荐,即便不打算延迟睡眠相位(注意 Seth Roberts 推荐不吃早餐,但这应适用于那些患有早醒或 ASPS 的人,而非入睡性失眠或 DSPS 患者)。不要被声称傍晚禁食不影响昼夜节律周期的研究所气馁。例如,Sensi 发现早晨的餐食似乎有利于碳水化合物代谢,且在研究范围内,用餐时机不影响昼夜节律周期(Sensi 和 Capani,198736)。禁食还有许多其他附加益处,包括更苗条的腰围。自己试一周,看看傍晚禁食对睡眠和健康的影响!更多阅读:Fasting。
-
增加压力:如果你持续因太晚醒来或在期望入睡时间不困而失败,继续在夜间睡眠两端加压。更多清晨运动、更亮的清晨光照、更长的傍晚保护区(无压力、电脑、光、电视等)。你的情况越难,你需要加得越多。选一个没有其他义务的日子做一次力竭运动马拉松(对你来说可能只是几小时快走),并把傍晚花在有助于提前入睡的不费力的放松活动上。你需要自己弄清楚什么运动强度能确保你不会过早醒来,以及你能承受多大的提前而不会让情况更糟。
-
应急:褪黑素:如果你的”保护区”已拉伸到极限,你可以考虑偶尔服用一粒褪黑素(例如在最佳入睡时间前 2 小时)。记住褪黑素会影响你的创造力和警觉性,不应常规使用。你可以在特别紧张或醒来时间特别晚的日子诉诸褪黑素。还要记住,大剂量褪黑素可能适得其反。我想你永远不应超过 3 毫克,然而你最好咨询睡眠专家以对剂量和时机做出最终决定。如果你持续在算法上失败,你应增加服用褪黑素的频率,而不是增加剂量!不要把大麻当作”褪黑素替代品”!如果你认为你的睡眠和大脑表现不受大麻影响,那你就错了!
-
应急:收音机定时器:由于算法应让你不靠闹钟睡眠,你不能指望每天在同一时间醒来。实际上,你的 DSPS 问题越大,预期变化越大。实现非常规律的醒来时间是需要练习的,一些好睡眠者经过多年重复习惯后才能做到。然而,如果你经常多睡超过一小时,或仍经历可怕的相位移动,你可以考虑偶尔使用温和的人为睡眠调节形式,如以最低音量打开闹钟收音机。这样温和的闹钟只是帮助你在清晨从最浅的睡眠阶段早醒,它不应剥夺你宝贵的 REM 睡眠,且绝对应设置在可能把你从深睡眠中唤醒的水平之下。你的目标始终是摆脱早晨的任何中断或干预。仅把它视为在注定包含小失败的初始适应期可能帮助你的临时措施。你可能听说过能读取你睡眠相位、帮助你最小化醒来痛苦的闹钟。我对这种解决方案并不热衷。概念上有道理,然而,它只是最小化可能切断睡眠关键部分的痛苦。在你电视最喜欢的频道上设置一个定时器,最低音量。如果它没能叫醒你,你就会知道你需要在算法的其他点上努力。试图为特定时刻醒来本身就足够有压力,可以作为许多人都相当有效的替代闹钟。一些科学家认为 ACTH 分泌被设定为确保及时醒来。我相信这种压缩的睡眠仍比被闹钟在更深阶段打断的睡眠好。至少,短睡眠后的自然醒来绝对比被闹钟在更深阶段打断的睡眠感觉好。总之,你应记住,你成功的决心也是算法的重要组成部分。决心可能不健康,可能影响你睡眠的激素谱,可能削弱你的免疫系统,然而,在不放弃现代生活方式的情况下平衡 DSPS 总会有一些残余的负面健康影响。只有回到古老的生活方式才能提供密闭的解决方案。
-
小睡:DSPS 中一个非常有争议的问题是小睡。许多专家会告诉你应完全避免小睡。我不同意。如果小睡太晚,可能引起相位移动。早的小睡(例如在清醒日第 6 小时)不会有这种效果。小睡将帮助你获得傍晚的生产力,尤其是如果清晨在阳光下早醒或清晨运动对你的早晨警觉性不太好。如果你持续无法实现 24 小时平衡周期,你可以逐渐把小睡移到更早时段。这将减少它们的长度和效率,但也将最小化它们的相位移动效应。只有在所有其他建议都失败时,你才应完全放弃小睡。如果你哀悼脑力敏锐度的丧失,记住我们并未演化成全天警觉和高效。100% 敏锐的头脑只是我们的欲望,它本身(在其他因素中)是 DSPS 问题的起因之一。见:Best nap timing。
-
咖啡因:睡眠专家常告诉你戒除咖啡因。然而,我恕不同意。一杯清晨咖啡或茶可能有有益效果。它对内稳态睡眠驱力的影响会增加你的警觉性,这本身具有相位移动力量。清晨咖啡因会帮助你前移相位!不过你应避免在一天较晚时段摄入咖啡因。睡前你将处于轻度戒断中,理论上这应增强你的内稳态睡眠驱力并允许稍早的入睡。这样,咖啡因可以帮助你在夜间睡眠两端实现相位前移。
-
酒精:睡前饮酒对你平衡周期的努力可能有灾难性影响。你的主要敌人之一将是过早入睡后的过早醒来。酒精会成倍增加那种风险。过早醒来会导致失眠、强大的相位移动、醒来困难、睡眠剥夺以及你所有努力的早期崩溃!西方文化通常容忍傍晚适度饮酒,而不容忍我们本应高效的早晨。我认为这种容忍应改变。清晨饮酒破坏思维的程度,与傍晚酒精可能通过破坏睡眠造成更大损害的程度相当。
-
压力:压力可能毁掉本算法中所有努力。只要可能,尽量把压力堆积在清晨,让一天的后半段始终免于担忧和焦虑。自然,持续数天的压力是健康高效生活的普遍敌人。如果这是你的问题,请瞥一眼本文的压力章节,看看是否能找到有用的提示。
-
近视:如果你近视,应考虑获得正确的处方。一些近视理论支持欠矫正(Rehm,198137),然而你也会发现相反的证据(Coghlan 和 Le Page,200238)。如果你倾向于欠矫正侧,考虑只在清晨使用正确处方。只有前 2-4 小时重要。你可能从物理课回忆起,到达视网膜的光量实际上会随焦距增加而减少。然而,聚焦良好的光更可能产生具有相位移动力量的强最大值。类似地,你也可以考虑在傍晚使用变暗的眼镜。自然,这些不推荐用于”近距离”应用(如看电脑屏幕),因为这可能加速近视进展。
-
电脑屏幕:有电脑屏幕滤镜和应用程序可以帮助你过滤相位移动的光频率,在傍晚调暗显示器。我从这些应用程序的用户那里听说,它们在 DSPS 战斗中有帮助。一如既往,存在这是安慰剂效应的风险。然而,尝试的成本不高。
-
困难:如果你持续经历相位移动,把午睡移到更早时间,或完全放弃(至少直到你重新获得平衡)。仅在最后手段时使用应急措施(褪黑素、收音机新闻)。如果这些最后手段解决方案持续必要,可能表明移动的自由睡眠周期比试图坚持”健康”稳定节律的损害更小。审查此列表上的所有建议。你是否都采用了?你是否展现了自律?如果是,它在大多数情况下应起作用。我见过的所有失败都来自相当站不住脚的借口和轻微的违反。我见过的所有成功都来自把睡眠作息当作宗教仪式来对待。如果所有其他都失败,请带上你的故事写信给我。
-
目标:在初期,经验不足、睡眠控制变量未对齐、睡眠债、缺乏信念等会使坚持本算法稍难。这对那些不需要早起上班的人来说尤为困难,他们的心思可能摇摆。早起值得这些努力吗?因此,在一个需要早起的重要活动前 3-4 周开始睡眠改革,在心理上可能有帮助。带着神清气爽的头脑参加重要活动的想法将增加遵守规则的动机。
-
创造力下降:一旦本算法对你起作用,你可能会失望地注意到,你的创造力不如那些你获得 8 小时充足睡眠的周末,或你凌晨 5 点入睡(取决于你的喜好)时那样旺盛。这是睡眠相位重置和相位前移措施不可避免的代价。我们演化来过 24 小时生活,首要关注的是生存、获取食物等。在逃离捕食者时,敏锐或创造性的头脑并不那么重要。当你在街上遇到狮子时,你必然处于逃跑的最佳状态。你创造性下降的程度可能与原始问题的程度,以及你用闹钟、安眠药和其他大脑健康敌人摆弄睡眠控制系统的时期长度成正比。
-
它有效! 记住,每个健康个体都能在农民的生活方式下坚持 24 小时周期。某些残障(如失明)可能使之不可能,但这些应是规则的例外。最坏的情况下,你可以尝试农民的生活方式,意识到它解决了你的睡眠问题,然后尝试把那个极端与本算法调和。在两者之间某处有一个适合你的最佳点,它不一定是不健康的。
-
终极治愈? 本章的标题不应让你认为你可以永久治愈 DSPS。它始终存在。你可以节制对现代生活诱惑的反应,回到平衡的睡眠模式作为回报。你的生活方式就是你的治愈。
结果:平衡的 24 小时睡眠周期!(Outcome: Balanced 24h sleep cycle!)
如果你使用 SleepChart,你可以在昼夜节律图中看到本算法的影响。下面给出了应用本算法的示例结果:
DSPS 中完美平衡的 24 小时周期。
在本例中:
- 绿线确定平衡 24 小时周期的条件(清醒时间 + 睡眠时间 = 24 小时)。
- 午睡在醒来后第 7 小时进行(左侧蓝色峰值),持续 1.2 小时(左侧紫色垂直线)。午睡用于部分补偿短的夜间睡眠。
- 最佳清醒日长度为 18.3 小时,由确定 24 小时睡眠-清醒周期的绿线与描绘平均睡眠时间的红色曲线的交点确定。更短的一天是可能的,但伴有入睡性失眠的风险。更长的一天将导致相位延迟(红线位于绿线之上的区域)。
- 最佳夜间睡眠长度为 5.7 小时(水平橙色线)。
- 该周期意味着夜间 5.7 小时睡眠和总计近 6.9 小时睡眠,这可能相当于单相睡眠中 9 小时的夜间睡眠(见:Optimum nap duration)。
总结(Summary)
在所给的算法中,你试图每天坚持最佳入睡时间和醒来时间。你在傍晚建立保护区以利于相位前移(最少的光、电脑、压力、兴奋等)。你在明亮的阳光下醒来,并用清晨运动在早晨前移相位。你只在清晨摄入咖啡因。你避免在傍晚饮酒。如果你小睡,你早睡。如果你的相位持续移动,你在清晨增加更多光照和运动,并延长傍晚的保护区。在紧急情况下,当你担心失步时,你可以偶尔在傍晚使用褪黑素,或在清晨使用轻柔的声音,作为对自由睡眠原则的最低有效偏离。
提前睡眠相位综合征(Advanced Sleep Phase Syndrome, ASPS)
提前睡眠相位综合征(ASPS)是 DSPS 的对立面。ASPS 患者在傍晚很早就非常困倦,并在夜里很早就醒来。他们的生物钟周期短于 24 小时,或在清晨很容易被重置(例如被压力)。ASPS 患者持续挣扎着清醒到合理的傍晚时段,睡得少、早醒,并在白天经历增加的疲倦。
ASPS 中的基因与生活方式(Genes and lifestyle in ASPS)
ASPS 常在家族中遗传,被称为家族性 ASPS 或 FASPS。一些可能导致 ASPS 的突变列在此表中。
典型的 DSPS 患者是青少年学生,典型的 ASPS 患者是退休者或耐压能力低的中年女性。年龄与睡眠相位障碍之间的联系可能与衰老本身有关,但也可能是伴随年龄而来的生活方式变化的结果。
值得注意的是,虽然我收到了数十份显示自由运行 DSPS 模式的 SleepChart 提交,我却不得不主动寻找能说明 ASPS 的提交。仅此一点就可以说明两组之间的人格和生活方式差异。是 DSPS 组在凌晨搜索睡眠问题的解决方案,最终到达 supermemo.com、下载 SleepChart,并开始记录睡眠以努力理解自己的睡眠模式。ASPS 组通常很早就上床,白天往往不够神清气爽,不会在网上寻求解决方案。我没有收到过哪怕一份睡眠时间在一天中越来越早的逆 ASPS 模式!
例子 1:ASPS 与物质滥用(Example 1: ASPS and substance abuse)
所给的 SleepChart 日志说明了一位绝经后、失业、有终身物质滥用史、目前处于戒断恢复期女性的稳定 ASPS 睡眠模式。没有药物,受试者挣扎着熬过下午 5 点。她常在凌晨 1-3 点醒来,发现无法重新入睡。她报告持续的疲倦。她睡眠问题的唯一解决方案似乎是由全科医生和/或精神科医生定期开的安眠药。这些药片也是苯二氮卓类和酒精成瘾恶性循环的一部分。
表面上看,日志似乎像完美健康睡眠的画面。然而,整个作息和睡眠相位是用精神活性药物鸡尾酒维持的。这个 ASPS 案例与看起来类似的完美睡眠案例的主要区别,是所说的全天持续疲倦。受试者报告她主要的关注点是”想办法熬过一天”,同时对抗疲倦并挣扎于随时可能回到成瘾的威胁。
各天睡眠长度的差异(好日子 8-9 小时,坏日子 0-3 小时)来自受试者不同天在不同家庭成员处睡觉的事实。其中一些被认为更好(导致更好的睡眠),另一些被认为压力更大。其中一个夜晚因家庭压力而失眠。这再次说明生活方式如何决定睡眠模式。
用药物维持的睡眠总是产生有限的认知收益。然而在这个案例中,效果确实惊人,认知表现与严重醉酒状态相当。药物作息因各种原因周期性变化,但不变地由早晨的提神药、傍晚的安眠药,以及一些控制”睡眠控制鸡尾酒”各种神经副作用的”额外”药物组成。各种药物互相干扰,产生一系列副作用,导致系统中可怕的混乱,以及长期后果,这反过来导致不可避免地滑向精神衰退,福祉、社会功能和长寿的大幅降低。对抗药物的半衰期导致它们的效果互相抵消,产生不可预测的综合后果。那么为什么这种药物恐怖会被容忍?对于一个有物质滥用史的失业者,对他/她的家庭和涉及的医生来说,今天任何类似常态的东西都优先于长期后果。自然,对几乎所有参与者来说,这种行尸走肉的现状优先于实际的醉酒,即便两者长期都注定摧毁大脑。脑电图发现表明,物质滥用和/或精神活性药物对中枢神经系统功能造成长期且很大程度上不可逆的变化。
与所有医疗干预一样,精神病学尤其受困于隧道视野,看不到个体生活和整体人口健康的大局。新药出现得太快,无法有效研究其长期后果,并受处方时尚的兴衰影响。随着巴比妥类让位于苯二氮卓类,苯二氮卓类让位于 Prozac,一个善意的精神科医生常被大量矛盾数据、永无止境的矛盾副作用清单,以及科学观察的灾祸——来自总是有多个理由对医生撒谎的患者的保证性错误数据——所困惑,使所有患者病史记录几乎变得无用。患者常更换医生以适应他们梦想的处方,或从不同来源寻求平行建议和矛盾处方。他们很少坚持药物时间和剂量。
为记录起见,截至撰写时,本例中使用的药物阵列是:
- 能够延长昼夜节律周期长度的抗抑郁药 文拉法辛(venlafaxine)(早上 8 点,半衰期 10 小时)
- 抗精神病药 阿立哌唑(aripiprazole)(早上 8 点,半衰期 75 小时)
- 抗焦虑苯二氮卓类 劳拉西泮(lorazepam)(晚上 6 点,2.5 毫克,半衰期 15 小时):使用了数年而非推荐的数周,尽管有各种副作用包括严重睡眠呼吸暂停;可能开处方的医生并不知情
- 抗精神病药 奥氮平(olanzapine)(晚上 8 点,20 毫克)
- 此外:抗惊厥药 拉莫三嗪(lamotrigine)(50 毫克)
截至撰写时,我无法确定这些是否由单一精神科医生开处,开处医生是否了解患者的完整病史。
例子 2:过度用药与白天困倦(Example 2: Overmedication and daytime drowsiness)
第二个例子显示了另一个严重用药的案例。56 岁的男性退休者依靠同样强效的药物鸡尾酒。在这个案例中,计划不周的不规律自由睡眠有助于揭示白天困倦的程度:睡眠事件规律地从清醒第 3 小时开始启动,第 7-9 小时有短而早的禁区,偏好 12-18 小时的短清醒日:
也许由于镇静剂的影响,睡眠事件的长度可能在一天中几乎任何时间达到相当于一整夜睡眠的程度。毋庸多言,受试者几乎无法进行认知运作,并抱怨永无止境的疲倦。本案例使用的药物:
- 早上 7-8 点之间:抗抑郁药 阿立哌唑(5 毫克)和 安非他酮(bupropion)(150 毫克)
- 上午 11:30 左右:抗精神病药 喹硫平(quetiapine)(25 毫克)
- 晚上 9 点左右:苯二氮卓类 氯硝西泮(clonazepam)(1 毫克),抗抑郁药 艾司西酞普兰(escitalopram)(40 毫克),拉莫三嗪(200 毫克),喹硫平(50 毫克)
- 此外:坦索罗辛(tamsulosin)以避免夜间尿失禁
相位移动图(Phase shift graph)
相位移动图可供患有 ASPS 或 DSPS 的自由睡眠者使用。此图显示相位移动的程度及其对一天中时间的依赖。该图可用于查看给定自然醒来时间的预期入睡时间:
蓝线显示给定醒来小时(横轴)那天的入睡时间(纵轴)。红线显示第二天的醒来小时(在 DSPS 中推迟 1-2 小时)。紫红色和灰线表示午睡时段。尽管红色醒来线从图表原点开始,它在较晚时段显示实质性的相位移动(DSPS)。从所给的示例图中可以读出,对于早上 7 点的醒来时间(横轴),蓝线指示的预期入睡时间是凌晨 1 点(纵轴),而最佳午睡时间在 15:00 到 16:00 之间。然而,如果醒来时间是上午 11 点,入睡时间可能只在次日凌晨 5 点。
记住! 每个个体都有自己独特的图。此外,图在工作时段和暑期休假时段看起来会不同。它受工作和家庭压力的影响。甚至你从一所房子搬到另一所,或改变气候带时也会变化。只有在你遵守自由睡眠时,该图才能准确反映你的节律。如果你使用闹钟,这张图将毫无意义!
睡眠相位综合征的相关因素(Correlates of sleep phase syndromes)
目前尚不清楚哪些是导致睡眠相位障碍的主要潜在生理因素。家族聚集表明基因可能影响昼夜节律周期的长度。生活方式会影响神经递质水平,并通过它们对睡眠相位的影响,也会影响昼夜节律时钟的周期。生活方式还影响授时因子的时机(例如 DSPS 中的深夜网上冲浪)。反过来,神经递质水平也可能选择特定的生活方式选择。年龄可能直接影响时钟回路,可能影响神经递质,也可能影响生活方式。最后,睡眠相位障碍会以不同方式影响情绪和神经递质水平,这取决于是否使用自由睡眠来治疗障碍,还是个体试图适应预先设定的理想睡眠作息。
昼夜节律睡眠障碍(CRSD)的子集与某些人格特征强烈相关,并可能有强烈的遗传背景。DSPS 在青少年中更普遍,而 ASPS 更常见于老龄化人口。女性在 ASPS 中占多数,而 DSPS 中男性比例略高(Sack 等,200739)。视力受损常因光对生物钟影响较小而导致 DSPS。
DSPS 在学生、程序员、 avid 读者、热情的艺术家、作家、电脑游戏成瘾者等中远更常见。可能有助于个体创造力的相同特征,也可能导致难以早睡的问题。ASPS 似乎更可能发生在那些生活缺乏强烈刺激(尤其是在傍晚)、面对更少新挑战、对工作或爱好不那么热情,或没有面临信息过载及相关压力的人身上。也许这就是 ASPS 在老年人中更普遍的原因。由于激素原因,其患病率在女性更年期前后也急剧上升。ASPS 倾向于家族聚集。已鉴定出一些基因与 FASPS(家族性 ASPS)有关(见:生物钟基因及影响时钟周期的突变(Golombek 和 Rosenstein,201030))。
DSPS/ASPS 与精神障碍之间存在复杂的关系。无法维持 24 小时睡眠-清醒周期的人中,25% 患有精神障碍(Hayakawa 等,199840)。一些精神障碍或处方药可能诱发 DSPS,同时 DSPS 反过来可能引起各种精神症状。一方面,DSPS 与躁狂人格之间可能存在联系。抗抑郁药倾向于延长生物钟周期(例如 氯吉灵(clorgyline)、丙咪嗪(imipramine))。另一方面,矛盾的是,DSPS 个体可能更容易患抑郁症(例如当遭受持续失眠、睡眠剥夺及由此产生的社会问题时)。Daniel Kripke 博士得出结论,DSPS 表型是家族性的,并与单相抑郁症相关(Kripke 等,200841)。然而,DSPS 在创造性个体中的流行表明,这些相关因素需要进一步研究。也许一些矛盾可以这样解释:DSPS 患者的心理状态很大程度上取决于他/她获得充足且时机恰当的睡眠的能力?因此在睡眠满足时(例如在自由睡眠作息下)更偏向躁狂侧,而在昼夜节律麻烦时(例如被迫早起作息时)更偏向抑郁侧?
类似地,耐压能力低、抑郁的个体更可能患 ASPS。同样,当他们被迫适应”正常”生活时,其抑郁症状倾向于减弱,这要么是因为更高的生产力感,要么是因为轻度睡眠剥夺对抗了抑郁。睡眠相位障碍与情绪障碍之间的因果关系是复杂的。理解它将大大有助于缓解睡眠问题日益升级的流行病。
婴儿睡眠(Baby sleep)
如何让婴儿睡好?(How to make babies sleep well?)
人们常说”我睡得像个婴儿?“一个笑话是,这意味着你每 2 小时醒来尖叫。确实,婴儿倾向于在夜里醒来,看似不高兴(除非立即用妈妈的乳房安抚)。这似乎不自然、不必要、令人担忧。然而,婴儿被设计成在夜里多次醒来进食。
网络上充斥着来自年轻父母、为他们宝宝和自己寻求好睡眠的各种建议。其中大量建议基于迷思和/或伪科学。如果建议包含”训练(train)“或”作息(schedule)“这样的词,你需要把怀疑度提高三倍!即便世界知名的儿科医生也过度强调”规律(routine)“而非实际的自然睡眠机制。与成年人一样,婴儿睡眠受内稳态和昼夜节律机制支配,任何试图覆盖这些的尝试都是徒劳且可能有害的。所有诸如摇晃宝宝、安静房间、喂食、轻声细语、音乐等规律都是受欢迎的,只要它们不是试图对宝宝强制执行睡眠作息。这些规律在诱导睡眠方面并不比所有奶奶的失眠建议更有效。内稳态和昼夜节律机制都不可训练(超出自然的相位移动等)。婴儿应按需睡眠(ad libitum),即只有当他们困倦、想睡时才睡。然而,理解他们的昼夜节律模式对辅助规律很有帮助。由于婴儿睡眠比成人睡眠更复杂,你可以用 SleepChart 看穿混乱。这可以作为指导。然而,观察困倦症状是最古老、最好的做法。
一些妈妈自豪地声称”我的宝宝能一觉睡到天亮”。然而,当分析实际睡眠日志或睡眠结构图时,这并非真实。很大一部分父母会不择手段让宝宝一觉睡到天亮。父母的方便和舒适常常优先于宝宝的健康。许多儿科医生对时间生物学规则相当无知,因为时间生物学在学校课程中不够突出。一些广受赞誉的方法简直令人恐惧。
“哭到睡着(cry-it-out)“法一定是受巴甫洛夫条件反射狗的方法启发。毫无疑问,长时间的哭泣和压力会抑制婴儿的发育。例如,如果长时间哭泣与后来的认知缺陷相关,至少在一定程度上与压力对发育的影响有关(Rao 等,200442)。归根结底,Ferber 的方法似乎服务于父母,而非孩子。任何父母的心都不应承受婴儿的哭泣,尤其是这完全是不必要的。
一觉睡到天亮(Sleeping throughout the night)
婴儿睡眠对齐成初步的昼夜节律模式大约需要 1-2 个月。这意味着最初新生儿对夜晚和白天区别不大! 与禁食相关的清醒的巩固先于母乳喂养节律的巩固,因为出生后前几周喂养需求很高(Odaa 等,200843)。
生命前 2 个月内稳态和昼夜节律睡眠偏好的示例昼夜节律图。未合并睡眠事件的平均长度(红线,左侧纵向数字线)从下午 4 点的 0.5 小时到晚上 9 点的 1.5 小时不等。对在傍晚早些时候启动睡眠(下午 5 点 6.2%)(蓝线,右侧纵向数字线)相对于早晨(早上 6-7 点低于 3%)有轻微昼夜偏好。使用各种睡眠合并方法(即将短时间间隔内连续的事件相加的方法),对傍晚睡眠的偏好可显示得更明显。
你的宝宝可能在白天和夜晚均等地重新分配睡眠事件。对于一个永远困倦的母亲,这可能导致错觉:宝宝顽固地倾向于白天睡觉,夜里却一直哭。难怪许多妈妈不断问:我做错了什么?、我怎么让宝宝在夜里睡? 简短的回答是:什么也没做(除非有破坏睡眠的器质性原因)!在 24 小时周期内以 1-4 小时短爆发形式睡眠对新生儿是正常的,对此无能为力!让宝宝在最初几个月一觉睡到天亮的努力是浪费时间! 此外,父母为达成该目标所尝试的任何事都可能对宝宝有害。
在下面的示例睡眠日志中,我们可以看到最初几个月的混乱如何慢慢合并成带有主要夜间睡眠事件和白天几次小睡的睡眠模式。大约 5 个月大时,相当一致的模式出现,大多数天有两次白天小睡。最后,大约一岁时,发展出类似成人的双相节律。突破通常出现在父母(未意识到合并过程)意识到宝宝不想早睡、并很快让孩子一天只睡一次的时候。这个阶段白天的多次小睡常是健康问题、错过某些夜间睡眠(例如因婴儿托儿早醒)、糟糕的”婴儿睡眠管理”(主要是对婴儿睡眠信号无反应),或生活方式变化(例如旅行、令人疲惫的游戏、见人等)的临时变化的结果。除非婴儿明确要求多次小睡,12-16 个月后单次午睡可能是一个相当安全的赌注。一些父母试图推着孩子整天保持清醒,以确保更扎实的夜间睡眠。然而,坚持孩子的自然偏好始终是更安全的选择。
生命第一年的示例自由睡眠日志。
有趣的是,在所给的图中,有一个 3 个月长的时期,婴儿倾向于很晚入睡。这种睡眠模式可能是对未来发育、心理或睡眠问题的令人担忧的前奏。然而在本例中,它也可能用冬季月份晨光重置影响较小来解释。对最初几个月睡眠的频谱分析表明,婴儿昼夜节律周期可能是四相的,组成频率随时间衰减,发展出典型的带有主要夜间和次要白天峰值的双相节律。
关于昼夜节律周期结晶的另一个例子,见这位一位父亲的努力,他绘制了宝宝睡眠中的规律。这张特定的图从宝宝主要是夜行性、但仍日间多相(第 4 个月)的阶段开始。大约第 10 个月,白天小睡合并,显示出四相模式,夜间”小睡”合并成单一的长夜间睡眠事件。最后,大约第 16 个月,孩子发展出清晰的双相模式。向双相模式的过渡可能因父母的决定(这常决定婴儿的睡眠时段)而有所延迟。
父母关于睡眠时机的决定将在很大程度上决定婴儿的睡眠模式。这就是为什么理解昼夜节律周期的自然发展并对自然婴儿睡眠信号做出反应,对健康婴儿睡眠至关重要!
健康昼夜节律周期的发展(Development of a healthy circadian cycle)
有两个主要因素会影响婴儿健康昼夜节律模式的发展:
- 暴露于自然的 24 小时光暗周期及所有相关的日常作息。
- 暴露于母亲的昼夜节律作息,由共同睡眠(co-sleeping)放大。
在许多动物中,昼夜节律周期的发展和初始驯化主要依赖于与哺乳母亲的互动(Rivkees 等,198844)。共同睡眠应有助于发展健康的昼夜节律周期。母亲在床上的存在以及母乳喂养都可以作为强大的线索。它们既充当 PRC 相关的也充当 PRC 无关的授时因子(见:相位响应曲线(PRC))。这意味着共同睡眠将影响睡眠相位以及夜间醒来的程度和总睡眠。除感官线索外,母乳喂养还扮演激素角色,因为母乳中色氨酸(tryptophan)的昼夜节律周期与儿童血液中褪黑素水平相关(由尿液中的 6-硫酸氧基褪黑素证明;Cubero 等,200545)。这自然让人质疑收集挤出的母乳供日后使用的做法。健康睡眠-清醒周期的发展自然也取决于母亲本人是否应用了适当的睡眠卫生原则。数百万儿童被迫独自在婴儿床里睡觉。这种做法在工业化国家如此普遍,我们可以有把握地得出结论:它不会不可逆地毁掉婴儿的昼夜节律周期,但在理论上,它可能是现代社会睡眠障碍流行病的根源。
除产后期外,母亲的昼夜节律周期在怀孕期间就已经对婴儿的大脑产生影响。这增加了孕期睡眠卫生的至高重要性。
夜间暴露于人工光是否会减慢或破坏昼夜节律周期发展过程,仍有争议。关于光对 SCN 发育影响的研究表明,有可能改变生物钟的节律性或对光的敏感度。发育过程中发生的变化可能终生影响时钟属性。然而,生活方式也可能逆转或放大这些变化。SCN 的发育在许多动物中被研究,结果各异。例如,大鼠 SCN 周期在子宫内发育(Altman 和 Bayer,197846),而负鼠 SCN 中的睡眠-清醒周期在出生后前 3 周发育(Rivkees 等,198844)。相位响应曲线的形状(位于睡眠相位障碍根源)可能实际上受发育期间照明条件的影响,至少在蟑螂中如此(Page,199147)。无论光和运动活动在婴儿中有什么影响,在我们了解更多之前,我们应始终致力于最小化夜间对人工光的暴露,并最小化其亮度(luminance)。
尽管常有相反的主张,夜间游戏实际上可能加速回到睡眠,只要重点放在身体而非情感上。然而,由于运动活动也能相位移动昼夜节律周期,应尽量减少按需的夜间游戏。
更多内容见:Polyphasic sleep in babies。
作为昼夜节律解决方案的共同睡眠(Co-sleeping as a circadian solution)
昼夜节律成熟的过程是神经性的,很大程度上超出父母控制。然而,妈妈和宝宝周期的驯化至关重要,可能决定该过程的最终结果。新生儿在 homeostatically 驱动下入睡,并主要由喂养需求唤醒。温度、饥饿、玩耍、光照、社交互动等因素只会增加画面的复杂性。由于有许多难以预测的因素影响睡眠需求,婴儿应始终按需睡眠。如果他们想玩,不应被拒绝。他们生活中所有的日程安排都应围绕睡眠进行。这基本上意味着新生儿好睡眠只有两个可行的解决方案:
-
推荐(McKenna 等,200548):妈妈和宝宝一起睡。妈妈需要学会适应整夜周期性的母乳喂养。妈妈的睡眠会受一些影响。然而,经过一些练习,母乳喂养可以最小程度地打扰,实际上对宝宝和妈妈都起到催眠作用。换言之,短暂的喂奶后,妈妈和宝宝都应迅速满足地入睡,因为任务完成得很好。即便昼夜节律周期的发展应无论如何进行,共同睡眠应辅助这一关键生物过程。与共同睡眠相关的婴儿安全问题必须事先解决(你可以在几分钟内用谷歌搜索;示例 1、示例 2)。即便一个强壮的一岁孩子也很容易被窒息。如果你相信你的强壮宝宝会尖叫或扭动,那不是真的!当其胸部被压缩或呼吸通道被阻塞时,婴儿几乎会瞬间失去力量,无法自卫!这就是为什么安全至关重要。对于一个健康、警觉的母亲,统计数据是好的。开始时一点焦虑是不可避免的,也许有助于发展健康安全的睡眠习惯。心理学家、行为主义者以及所有思想开放的儿科医生都会承认,夜间纽带对婴儿大脑发育极好。对妈妈和孩子都是完美的公式。
-
应急:父母借助母乳轮流照看。可能出于健康原因,妈妈和宝宝应暂时分开睡。在这种情况下,如果父母适应一种借助时间疗法的轮班工作变体,双方仍能获得极佳睡眠。例如,一位父母在时间生物学上适应晚上 7 点 - 凌晨 1 点时段,另一位承担凌晨 1 点 - 早上 7 点班。在妈妈睡觉期间,可以使用事先挤出的母乳(或配方奶)代替母乳喂养,以避免对妈妈睡眠的重大打扰(记住,母乳特性也沿睡眠-清醒周期变化)。如果该解决方案因任何原因不可行,父母可能需要在以下两者之间选择:
- 他们自己的好睡眠,以及
- 雇佣夜间保姆。
毋庸多言,婴儿的睡眠在此绝不应受到威胁。睡眠对成人健康至关重要。然而,对婴儿来说,这简直是生死攸关的事,因为睡眠破坏会影响许多新生儿死亡率的原因。
最佳喂养时机(Best timing for feeding)
除非有资格的儿科医生因特定健康原因特别指示,“按钟喂食”应被禁止。永远不要为了喂养叫醒宝宝!按需喂养是宝宝的最佳选择!
为人父母的弥天大罪之一是过度喂养!
- 容易观察到,匆忙吸下一大剂配方奶后,婴儿因与实际困倦无关的原因而昏昏欲睡。把宝宝喂到睡可能很诱人。这种做法对宝宝健康有害,不会帮助他/她整夜睡眠。它只可能辅助短暂睡眠,清除内稳态困倦,使宝宝比喂养前更不困。
- 过度喂养的另一个原因是妈妈的方便。为了获得更多不被打扰的睡眠,妈妈会在夜间或夜间喂养前给宝宝额外的配方奶。同样,父母的方便可能对孩子有害!
孩子自己的床(Child’s own bed)
美国儿科学会令人惊讶地反对共同睡眠!许多妈妈会发誓她们的宝宝可以独自在婴儿床里睡觉而没有任何痛苦。然而大多数行为主义者和人类学家会同意:妈妈和宝宝应密切接触睡眠,正如人类千年来、筑巢哺乳动物数百万年来所实践的那样。出于健康或安全理由可能有例外。然而,对普通妈妈来说,与宝宝同睡应是一种乐趣、特权和责任。年轻妈妈只需阅读基本的安全措施。对宝宝来说,与妈妈同睡应是基本人权!许多儿科医生、护士、助产士和”老派”奶奶仍会坚持宝宝应睡在自己的床上,出于安全、纪律或便利原因。你可能听到医疗专业人士建议:“不要奖励宝宝!你是在自找麻烦。” 鉴于婴儿奖励系统相当精调地满足其生物需求,这令人惊讶。一个控制机制被现代生活方式搞乱的普通成人常常应被拒绝奖励(一个甜甜圈、一杯晨间威士忌等)。然而,所有自然奖励对婴儿应被认为在生物学上有利。至于安全问题,宝宝确实在与妈妈同睡时、在软床上、因酒精等而死亡,独自睡觉时也是如此。简单的预防措施能极大地降低可怕事故的风险。我们应从出生起训练孩子独自睡觉的想法难以根除。人们需要仔细审视母乳喂养和婴儿睡眠的生物学,才能迅速意识到在那个阶段训练无异于残忍(除了宝宝不反对独自在婴儿床或妈妈离开共同床的情况和时刻)。一个小婴儿基本上是一台喂养、生长和学习机器。其所有先天反射都旨在确保安全、生长和大脑发育。寻乳、吸吮和母乳喂养的反射是小婴儿最强的驱力之一。一整套脑中心参与其中,这些中心与睡眠中心以及幸福感和愉悦感之间存在密切关系。尽管所涉机制的复杂性远未被我们的理解揭示,拒绝宝宝妈妈的乳房和亲密接触注定会有长期的发育后果。打个比方,你可以把自己置于宝宝的处境,尝试裸睡在冰冷的水泥地上,整夜大声播放恐怖电影。我可能在这里过于戏剧化。如果宝宝自己不声不响地入睡,其对亲密的渴望显然不像这里描述的那样强烈。然而,当宝宝的生长受到威胁时,你应始终偏向安全侧。这就是为什么假设最坏情况是有意义的。如果研究人员随着时间的推移发现,需要在婴儿的睡眠倾向双过程模型中扩展一个涉及母乳喂养的因素,我不会惊讶。已经表明,乳房是催眠剂。如果它作为婴儿内稳态睡眠倾向的组成部分甚至触发器起作用,我不会惊讶,尤其是在昼夜节律周期尚未完全表达的时期。毋庸多言,婴儿比成人更需要睡眠。年轻大脑中神经生长、网络重塑和学习的程度是惊人的。NREM 和 REM 睡眠成分在此过程中都不可或缺。改变睡眠结构将影响神经发生(Stryker 等,200130)。换言之,睡眠前或睡眠中任何形式的压力都会影响婴儿的大脑生长。这就是为什么婴儿的睡眠应是最高保护区域。训练独自睡觉可以等待。早期断奶的小猪已被证明其海马体受损,导致人格变化、对探索的恐惧和低耐压能力。
许多父母反对亲密育儿(attachment parenting),认为它在时间上太昂贵。很难与需要在养家和正确行为方法之间选择的人争论。在这里,现代生活方式再次以长期后果的恶性循环入侵人类生物学:用流水线方法养大的婴儿在情感和智力上更可能无法应对调和科技进步与社会进步与人体和大脑需求的挑战。更多内容见:Is sleeping with my baby safe?(McKenna,199549)和 The Science of Attachment Parenting。
妈妈怎么办?(What about the mom?)
共同睡眠对妈妈好吗?我相信每个人都应品尝自由睡眠的福祉。然而,在婴儿睡眠受到威胁时,我的一些良好睡眠倡导需要例外。每位母亲都配备了潜在或显性的本能,应使共同睡眠的体验愉悦。自然,在现代世界,压力、手机、电视、互联网、激烈竞争等因素会使之非常困难。如果一位妈妈声称”我讨厌母乳喂养”或”我无法与宝宝同睡”,她应从彻底审视自己的生活开始。如果家庭足够免受室外风暴的侵袭,母乳喂养和共同睡眠会非常有回报。
为什么婴儿睡这么多?(Why babies sleep so much?)
婴儿睡这么多是因为他们学得多,还是因为他们睡得多所以学得多?婴儿睡这么多是因为他们的大脑被设计成在生命的最初几个月这样做。他们确实学得多,学习确实增加睡眠需求,但这不是主要的调节因素。婴儿的睡眠控制系统运作方式不同,你可能无法通过让他们学得更少来让他们睡得更少。另一方面,长时间的睡眠被用于重组大脑中的神经网络。简而言之,睡眠帮助学习,学习诱导睡眠,但整个睡眠序列是年轻睡眠控制系统基因编程属性的直接结果。考虑到婴儿将约 50% 的睡眠时间花在 REM 阶段(相比之下成人约 20%),有一种理论认为,即便婴儿白天在探索周围环境时学得不多,这一功能也由 REM 睡眠的探索功能填补,帮助他们发现已学事物的新属性和规则。
你常听说新生儿大部分时间都在睡觉。实际测量可能表明,婴儿更可能只睡掉他们一半的日子,而其中只有一小部分处于较深的 NREM 睡眠阶段。

生命第一个月的示例 SleepChart 睡眠日志。所有睡眠区块的总和显示在右侧,平均每天略高于 10 小时,并有显著的日间波动,反映了丰富的内稳态输入变化(如散步、家庭访问、尿布疹、配方奶补充等)的影响。
结论:婴儿睡眠的完美公式(Conclusion: Perfect formula for baby sleep)
婴儿按需睡眠、按需喂养时睡得最好! 任何对这些内稳态调节模式的人为干预都应被认为可能有害。发现规律并额外关注宝宝在偏好喂养和睡眠时段的信号是有帮助的,然而,是宝宝的需求决定实际时机。由于睡眠对生长大脑中神经结构的发育至关重要(Stryker 等,200130),任何形式的干预和人为控制从长期看都应被认为可能有害。
如果你想让宝宝聪明健康,让它按需玩耍、按需喂养、按需睡眠。
失眠(Insomnia)
失眠是入睡或保持睡眠的困难。心理生理性失眠常持续数年,并对人的生活造成难以估量的损害。那些绝望到去看医生的人常被开安眠药,这通常不比失眠本身好多少!好消息是,在大多数情况下,失眠可以通过下面描述的睡眠相位调整轻松治愈。坏消息是,这种调整可能与期望的工作或学校作息不兼容。
工业化国家一半的人口在入睡方面有问题!这被称为入睡性失眠(sleep onset insomnia)。除各种潜在的器质性原因外,绝大多数入睡性失眠病例源于无法驯化睡眠时段以匹配期望的醒来时间。
换言之,大多数在其他方面健康、晚上无法入睡的人,苦于两个主要因素的组合:
- 相对于其自然昼夜节律周期过早入睡
- 承受相关的压力:如果我不立即入睡,我早上会完全废掉
如果这些人被允许想睡多久睡多久,并且只有真正疲倦时(也许晚 2-5 小时)才入睡,在绝大多数情况下问题将不存在!一些科学家把失眠称为去唤醒(de-arousal)过程的抑制。在睡眠相位问题中,去唤醒的问题不存在!去唤醒正确进行,只是在一个较晚的相位进行。
对年轻的求学人群,睡眠相位问题是失眠最常见的原因。对需要早起上学的学生,他们的睡眠相位常被定位得太晚,相对于期望的醒来时刻。换言之,最佳睡眠时间来得太晚,困倦来得太晚,自然醒来也相应地晚。这样的学生在晚睡时会一直与睡眠剥夺斗争,或在早睡时有某种程度的失眠。在这个意义上,存在物理/生物的潜在原因。然而,由于睡眠剥夺相当不愉快,学生可能试图早睡(以确保夜晚足够长),却因昼夜节律时段过早而无法入睡。如果这种情况反复发生,心理因素可能加重原有的失眠问题。反复的睡眠剥夺会产生对不及时入睡、让事情更糟的恐惧。换言之,在绝大多数情况下,问题既是生物学的又是心理学的。唯一真正的解药是晚睡晚起,从而上学迟到(在教育者一方给予一些理解的情况下,这几乎肯定是较小的恶)。唯一自然的半解药是尽可能精确地测量最佳入睡时间,并每天一丝不苟地坚持那个时间。那个最佳时间是大致提供 95% 或以上把握睡眠潜伏期少于 10-15 分钟(即不超过一刻钟的辗转反侧)的最早时间。这个最佳时间通常只提供 4-6 小时的睡眠。然而,这种睡眠最可能是这种条件下可达到的最佳质量睡眠。自然,受影响者每天会承受一定程度的睡眠剥夺。这仍比徒劳的辗转反侧、浪费时间以及与失眠相关的浅睡要好。如果你患有入睡性失眠,并怀疑它可能由 DSPS 引起,你可以研究额外的疗法,如清晨运动、强烈的清晨光照、傍晚褪黑素,以及激进的解决方案,如……19:00 之后放弃用电。
另一种失眠,夜间醒来(nocturnal awakening),也常与在错误时间入睡有关。需要比生物钟指示更早起床的人,常试图通过早睡来弥补短暂的夜晚。如果他们成功入睡,常会经历过早醒来,这几乎总伴随着重新启动睡眠的困难。如果这些人被允许只在真正疲倦时(也许晚 2-5 小时)才入睡,问题很可能不存在!
如果你经常在夜里醒来,应识别并消除可能的原因,尤其是如果你醒来时显得疲倦。原因和诊断方法太多,无法一一描述。然而,你应始终从最简单的开始:昼夜节律相位问题。简单地说,你的睡眠时机可能错了。睡眠的分段是过早入睡的典型症状。如果你健康,在自由睡眠中,你很少会在夜里醒来,即便醒来也会很快重新入睡,如果不能,原因会相当明显,如压力、噪音、口渴、寒冷、膀胱满等。然而,如果你试图调节睡眠时机,睡眠分段(即被打断的睡眠)将是频繁的结果。把睡眠稍微提前或推迟(例如每天 15-25 分钟)而不产生这种负面结果是可能的。然而,一旦你试图推得太狠(例如每天超过一小时),分段几乎不可避免。如果你向后推(即越来越早入睡),你很可能在夜里早醒,即在昼夜节律低谷确保深睡之前。另一方面,如果你向前推(即越来越晚入睡),你的昼夜节律低谷将在你完成睡眠周期之前结束。结果,你常会比预期更早醒来。如果这种醒来发生得很早(当你向前推得很狠时),你会足够疲倦地再次入睡。换言之,无论你向哪个方向推睡眠,它都不会与昼夜节律正确对齐。然后你会在睡眠周期的早端或晚端醒来,取决于错位发生在哪一端。在绝大多数情况下,醒来问题可以通过在身体呼唤时入睡来解决。
大多数入睡性失眠和夜间醒来案例的解决方案是:只有当你真正困倦时才入睡! 令人惊讶的是,大多数人不愿倾听自己的身体。许多人在傍晚与困倦斗争以获得更多生活,另一些人则在最佳入睡时间很久之前强迫自己上床,然后辗转反侧几个小时。这种过早的上床是失眠流行病的根源(尽管官方数据把昼夜节律障碍只列在失眠原因的 10%)。唯一明智且健康的入睡时间是当你感觉开始困倦时。如果这个自然时间离谱地晚,见:Curing DSPS and insomnia。
早醒也是许多人的问题。这些人可能患有 ASPS 综合征。在他们的情况下,早睡常是足够的疗法。如果你碰巧在清晨早醒,进一步的睡眠决定可能应基于你认为自己能多快入睡。如果你认为睡眠不会很快到来,起床做点工作肯定是更好的选择。这样你将在三个方面受益:
- 更快地感到疲倦和困倦(工作更可能让你再次疲倦)
- 你不会因尽快重新入睡而焦虑
- 你不会在徒劳的辗转反侧上浪费时间
失眠与电(Insomnia vs. electricity)
自人工照明出现以来,失眠已达流行病程度。看看失眠的终生成本如何与工业化程度相匹配。
无数据
少于 25
25–30.25
30.25–36
36–41.5
41.5–47
47–52.5
52.5–58
58–63.5
63.5–69
69–74.5
74.5–80
多于 80
如何更快入睡?(How to fall asleep faster?)
关于入睡性失眠有冗长的书籍,人们使用无数的技巧来确保”按时”入睡。可悲的真相是,所有这些技巧都在与不可避免的事作斗争:自然睡眠机制。它们基于在大脑根本不想减速时让它减速。然而这些技巧往往通过增加睡眠者的压力而使失眠问题恶化:这么多努力,这么多技巧,仍然不起作用……我可能只能永远与这个灾祸共存!
如果你遵循传统的失眠建议(见一个示例),你会很快意识到大多数措施在开始时效果很好,然后当安慰剂效应消退时,你又回到原点。一些酒店为时差旅客提供每晚 1000 美元的失眠治愈服务。所有这些服务都是对时间和金钱的巨大浪费。没有相位调整,失眠将持续,只能被暂时掩盖。
以下是一些你可能从睡眠”专家”或奶奶那里得到的典型无效建议:
- 无效!数羊——这只是试图减慢大脑中的神经元放电。这常会奏效,但存在 20-60 分钟睡眠后醒来的增加风险。结果,早睡的机会可能永远丧失。放电减慢可以自然到来。然而,它只能在正确的昼夜节律时间到来,这也许比你想要的晚 2-5 小时。
- 无效!思想分散——通过试图”什么都不想”,你确实可以增加入睡的机会;然而,这只在上升的昼夜节律睡眠倾向斜坡之前的短时期有价值。如果你过早尝试,你很可能在浅睡中徘徊,有相当大的自发醒来机会。是昼夜节律困倦在维持你的睡眠能力。
- 无效!减少睡眠——这在重置昼夜节律周期方面有一定力量,然而,你会更疲倦,认知能力下降,失眠者的压力因素增加三倍,最重要的是……你的创造性大脑不会从最后富含 REM 的几小时睡眠中受益!减少睡眠对那些患有 DSPS 且无法自由睡眠的人来说可能是不可避免的解决方案。
- 有害!安眠药(尤其是苯二氮卓类)——药物改变睡眠生理,可能影响睡眠质量。由于它们对修改实际昼夜节律周期影响甚微,它们可能迅速成为严重成瘾。此外,它们影响短期记忆和反应时间。安眠药是创造型个人的主要敌人!
- 无效!温水浴、安静房间、仪式、一杯可可等——所有这些都旨在减少压力因素和减慢大脑。然而,如果你相对于昼夜节律周期过早尝试,同样不会奏效。
- 无效!喝牛奶或吃火鸡(色氨酸用于合成褪黑素)——这又是一种温和减慢的方法,效果微乎其微。
- 无效!服用维生素——维生素或矿物质是健康出现问题时标准的万能借口。对许多人来说,第一个问题是*“也许我维生素摄入不足?”* 加倍推荐剂量是常见后果。健康饮食很少需要补充!
- 无效!不小睡——时机恰当的短小睡对昼夜节律周期影响甚微,不小睡可能只在傍晚警觉性的代价上产生残余的内稳态影响。它可能略有帮助,但不会解决问题。只有在昼夜节律严重错位、身体开始失去时间感、昼夜节律变量开始不同步振荡时才应避免小睡。
- 无效!避免咖啡因——当在清晨适量摄入时,咖啡因对睡眠质量影响应该可以忽略。当你真正疲倦时,很难用咖啡阻止睡眠。自然,戒除成瘾(逐渐!)会略有帮助且无害。然而,它永远不会是解决方案!
- 无效!草药、冥想、瑜伽、芳香疗法、针灸、洋甘菊、放松、按摩、镁……——……这个列表几乎无穷无尽!
对高效人群来说,在身体不想睡的时候试图入睡,能产生的时间浪费无出其右。不要听信仅基于傍晚减慢方法或让你精神或身体疲倦的睡眠建议!在大脑自己减慢之前不要上床!只有当你真正困倦时才上床!
本节标题提出的问题是有意挑衅的。你无法更快入睡,但你可以快速入睡。 你需要做的只是等待正确的时机。与其试图更快入睡,不如晚些入睡,快速入睡。
基于相位移动的失眠(Phase-shift based insomnia)
在大多数情况下,失眠的真正罪魁祸首是你的工作时间与昼夜节律之间的关系!这被相关的压力因素大大放大。对许多人来说,失眠产生了一个必须与之共活的无法解决的恶性循环。然而,每个有机会使用弹性工作制或远程办公的人都应意识到,这些的最大好处可能来自因更好睡眠(符合自然身体节律)而带来的生产力提升。
在 DSPS 患者中,需要特定程度的”早晨痛苦”来充分重置时钟。在痛苦足以维持规律作息的均衡状态下,整夜睡眠被大幅削减,导致日常睡眠亏欠和日常与疲倦的斗争。在这种情况下,最好只在预期入睡时间前不久才入睡!这样你可以一方面减少压力,另一方面帮助你的内稳态组分(让自己为睡眠而疲倦)。
Kripke 博士(见关于短睡眠的批判章节)说:“人们普遍需要 8 小时睡眠的错误信念是失眠的常见原因之一。在床上花更少时间是许多失眠患者的重要解决方案。” 这一说法只部分正确。确实,试图通过早睡来获得 8 小时睡眠会适得其反。在正确相位启动时少睡,比过早强求额外小时更好。然而,关键不在于少睡,而在于在正确的时间睡!如果你在正确相位入睡且不需要早起,你实际上可以获得 8 小时睡眠而零失眠风险。
如果你无法自由睡眠,让你的”早晨痛苦”尽可能规律以达到均衡状态。一旦你知道均衡点,坚持你标准的入睡时间。早晨痛苦的解决方案应只作为最后手段!
周末的问题仍然存在。许多人在周末补回失去的睡眠。这自然使系统失衡,导致”周一早晨忧郁症”。在周末睡懒觉时,你应权衡利弊:
- 一方面,你把睡眠周期驯化到更晚时段,使更难坚持”痛苦均衡”;
- 另一方面,这是你获得高质量睡眠的唯一机会。
周末困境没有简单答案!如果你想把睡眠对学习、技能和经验的影响最大化,你需要量化因从未真正获得足够睡眠而损失多少(损失可能巨大!),以及因周末偏离痛苦均衡从而使一周早期睡眠干扰增加三倍而损失多少。
对健康人来说,持续失眠或与工作作息相关的睡眠剥夺最有效的解决方案是自由睡眠!
自由睡眠定义简单,但对初学者来说执行起来有点难。它常与个人对睡眠时机的期望和需求冲突。如果失眠被你上床后不超过 3-5 分钟(不用药物)入睡所取代,你就知道自由睡眠对你起作用了。
对健康人来说,入睡时间不应超过 5 分钟!
如果你在自由睡眠上成功,并发现只要在自然时间入睡就能 5 分钟入睡,你可能会发现一个新问题:你可能倾向于每天越来越晚醒来。如果是这种情况,阅读:Curing DSPS and insomnia。
嗜睡(Hypersomnia)
嗜睡(hypersomnia)是在获得生理上充足睡眠的条件下的过度困倦。嗜睡可能与严重健康问题有关。然而,如果你持续与困倦作斗争,你的问题不一定有严重的器质性原因。有一种简单的家庭诊断方法可以帮助你排除一个常见且较轻的原因:相位移动障碍。尝试自由睡眠几周。相位调整常常能解决持续的疲倦!非常常见的是,相对于昼夜节律困倦过早启动的睡眠会持续很长,并矛盾地导致早晨不神清气爽的感觉。如果主观昼夜节律夜晚时段与实际醒来时间重叠,你可能会经历压倒性的困倦,却无法入睡超过 20-30 分钟,醒来后仍不神清气爽。在这种情况下,一桶桶咖啡也无济于事。如果在 1-2 周自由睡眠后没有注意到睡眠质量的显著改善,你可能患有需要专业咨询的潜在健康问题。见 WebMD 的 Sleep and Hypersomnia。质量差的睡眠的常见原因是阻塞性睡眠呼吸暂停(OSA)。OSA 由睡眠中的呼吸困难引起(见下一节)。
睡眠呼吸暂停(Sleep apnea)
睡眠呼吸暂停是睡眠中的呼吸问题。在所有遵守所有睡眠卫生规则(尤其是在自由睡眠中)仍获得不令人神清气爽睡眠的案例中,需要排除睡眠呼吸暂停。初步的家庭诊断可由床伴或通过录制自己睡眠的视频做出。所有呼吸暂停或严重打鼾都应令人担忧并咨询睡眠专家。最常见的睡眠呼吸暂停类型是阻塞性睡眠呼吸暂停(obstructive sleep apnea)(OSA),影响高达 10% 的男性人口(女性约为一半频率)。OSA 涉及喉部和舌部区域肌肉张力丧失,这些结构在睡眠中倾向于塌陷并阻塞气流。结果,患者会暂时醒来(常常一夜上百次)而未完成自然的 NREM-REM 周期。OSA 患者醒来时感觉不神清气爽。你也可以在睡觉时给自己录像。OSA 最常影响肥胖和严重打鼾的男性。网上有多个 OSA 支持站点(包括打鼾患者的录音和呼吸中断的典型迹象)。睡眠呼吸暂停需要紧急治疗。它在短期内(通过对睡眠质量的影响)和长期内(通过对大脑生长和衰老的负面影响)削弱人的认知能力。如果有人告诉你你严重打鼾,不要把它当作自然的事:“我叔叔也像拖拉机一样打鼾,似乎没事。” 所有打鼾案例都应调查。打鼾可能是你的大脑在夜间睡眠中得不到所需的第一迹象!睡眠呼吸暂停还可能导致心血管疾病、抑郁症和一系列负面健康后果。对抗睡眠呼吸暂停最好的自然武器之一是通过健康饮食和运动相结合来减肥。运动本身也可能有帮助,只要仔细选择以确保不会加剧打鼾(例如通过鼻塞)。
打盹(Napping)
打盹是好事(Napping is good)
自然的睡眠-清醒周期让你在中午感觉不那么警觉。这个时段可以用脑电图(EEG)测量轻松可视化。在许多热带、亚热带和地中海国家,这是午睡(siesta)的时间。丰盛的一餐会放大警觉性的下降,而短暂的小睡可能迅速恢复完全的警觉性。然而,工业化国家似乎并未准备好采纳午后小睡这一健康习惯。恰恰相反,当墨西哥议会辩论法定小睡法案时,边境以北的政客和喜剧演员对”懒惰的拉美人”好一番嘲笑。英国的 Siesta Awareness 在中国一篇显示糖尿病患者小睡更多的文章发表后,突然取消了他们的 2009 年国家午睡日。迷思比比皆是。打盹是聪明的,然而打盹者常被认为懒惰或软弱。自助大师 Tony Robbins 提供了一个典型的、误导性的”打起精神来”建议:用俯卧撑替代小睡的冲动。俯卧撑会改善血液循环并提高儿茶酚胺水平,让你一时感觉更警觉。然而,只有小睡才能为你的认知能力提供真正的神经提升。小睡胜过运动,小睡胜过咖啡因,小睡不可替代。
打盹与演化(Napping and evolution)
关于中午警觉性低谷的演化目的,理论不多。大多数人认为,人类如同所有其他高度发达的热带动物一样,发展出午睡习惯是为了应对正午的炎热。这一解释也有一些文化背景,因为在温和和寒冷气候中小睡远不那么流行。然而,警觉性低谷可以在几分钟内通过短暂小睡解决,这能让我们在正午炎热结束前很久就重新活跃。
另一种解释是,警觉性低谷是可能曾是人类祖先特征的多相睡眠模式的无用残留。许多动物和新生儿在一天中睡很多次,这似乎有利于优化记忆回路。然而,把睡眠整合成单一的夜间休息时段可能也提供了某种演化优势。早期人类在夜间的狩猎和采集活动可能效率较低,因此把夜晚用于记忆优化可能是有利的。可能,睡眠的整合经历了从多相睡眠、经过双相睡眠、到现代人半单相睡眠的逐步过程。实际上,类似的整合可以随着年龄增长观察到。到成年时,我们或多或少是单相的,有一个明显的警觉性低谷,可以通过短暂小睡解决。临近退休时,我们似乎又倾向于变成双相。这可能是工作人群被迫压制其双相倾向的结果。我们在整个生命周期中保持强烈的双相本性,单相模式很大程度上是由工业化强加的。
当我查看用 SuperMemo 收集的学习表现数据时,我看到一天中认知能力的内稳态下降足够陡峭,以至于可以提供一个替代解释:小睡在认知上有益,但还没重要到需要昼夜节律的全力支持。因此,我们发展出一个折中的睡眠系统:确保必不可少的、完整的夜间睡眠,以及一个可选的、中午警觉性提升的窗口。由于睡眠驱力的昼夜节律组分与睡眠的某些生理功能相关,一个以内稳态小睡为基础的系统可能同样不那么有益。至于内稳态警觉性下降的速度,它可能是所涉网络固有的,可能取决于能量储备、神经递质供应、所涉网络的规模等,还应取决于使用程度。脑力付出越重,认知表现下降越快。换言之,对现在这样的大脑,对沉重的脑力负荷,较慢的内稳态下降可能根本在物理上不可能。中午小睡的时机来自这样一个事实:把一天分成精确的两半能最大化总体警觉性。在这里,正午的热带炎热可能确实提供了额外的演化激励。
小睡与脑力(Naps and brain power)
小睡科学之父 D. F. Dinges 博士花多年时间研究工作场所的警觉性问题,并证明在警觉性可能关乎生死的职业中,小睡有实质性的好处。他的研究表明小睡带来显著的警觉性提升(Dinges,198923)。他还注意到,规律小睡者的小睡对夜间睡眠的影响相对较小:
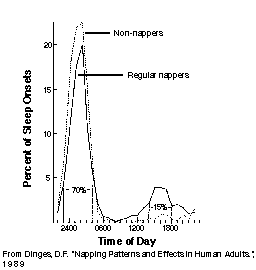
然而,当 Matthew Walker 博士发表其证明小睡对认知价值的研究(Walker 和 Stickgold,200550;Walker 和 Nishida,200751)时,Derk-Jan Dijk 教授却令人惊讶地评论:“没有明确证据表明白天小睡相对 24 小时只睡一次有明显优势(……)虽然研究中报告的大脑效应可能在实验室环境下被捕捉到,但在’真实世界’中画面变得更加模糊”。如今,你可以用 SuperMemo 自己测量小睡的好处。比较小睡者和不小睡者的回忆图,我们可以清楚地看到不小睡者在清醒日的下半天以半蒸汽状态运行(见:Biphasic nature of human sleep)。Walker 博士(其观点被后来的研究证实)令人信服地说:“就好像你海马体里的电子邮件收件箱满了,在你睡眠并清理掉所有这些事实邮件之前,你不会再收到任何邮件”。听听虔诚的小睡者温斯顿·丘吉尔(Winston Churchill)的话:“你一天当两天用!” 小睡的价值与你工作对大脑和思维质量的依赖程度成正比。
小睡还是不小睡?小睡!(To nap or not to nap? Nap!)
以下是午后小睡利弊的简短总结:
好处:
- 富含 NREM 睡眠的午睡小睡,会导致警觉性的显著提升,这将深受创造型职业人士赞赏。通过各种测量,这种提升可高达 50%!
- 如 SleepChart 所示,小睡改善下半天的回忆和记忆巩固。
- 时机恰当的小睡可帮助对抗睡眠剥夺。一些人甚至终身更喜欢分两段各 3-4 小时睡眠!
- 小睡降低血压、改善心血管健康,可能因此延长寿命!
坏处:
- 时机不当(过晚)的小睡可能加重失眠。这就是为什么那么多睡眠专家反向推荐避免小睡。然而,恰当的时机可以缓解这个问题。
- 时机不当(过晚)的小睡可能导致睡眠惰性。这可以通过恰当的小睡时机轻松解决。
- 小睡需要良好的休息条件、有利于小睡的文化以及扎实的小睡技能和习惯。在工业化国家,这些都可能难以获得。
如果你曾犹豫,小睡还是不小睡,就在恰当的时间小睡一次,看看它如何影响你的生活。如果你醒来昏沉,记住小睡也是一门艺术。阅读关于最佳小睡时机的内容。很可能,小睡会成为你深爱的、能提高你生产力的习惯。许多伟大的头脑习惯性地小睡。除了丘吉尔,著名的小睡者还包括拿破仑、比尔·克林顿和 J. F. 肯尼迪。有趣的是,这一群体还包括著名的长睡眠者阿尔伯特·爱因斯坦和著名的短睡眠者托马斯·爱迪生。甚至比尔·盖茨在他创造性的编程年代也喜欢在桌子底下小睡。
企业界的小睡(Napping in the corporate world)
美国越来越多的公司已经决定从咖啡休息时间转向小睡休息时间,配有专为小睡者设计的隔间。未来,这一趋势可能变得更加突出,因为咖啡因在对抗疲劳方面不及小睡的几分之一。出于神经原因,咖啡、甜甜圈、俯卧撑及其他方法加在一起,在精神恢复方面永远无法像小睡那样高效。同时,我们的社会强烈地向信息处理(其中警觉性是生产力的核心)漂移。当生产力进入方程,企业肯定会利用关于小睡的最新研究。
重要! 不要把午睡的健康概念与非常不健康的多相睡眠(polyphasic sleep)想法混淆。
小睡规则手册(Napping rulebook)
- 不要使用闹钟!与普遍看法相反,安排得当的小睡不会导致睡眠惰性(除非你严重睡眠剥夺)。闹钟会严重削弱小睡的价值!
- 精确测量自然醒来与小睡之间的最佳时段长度,以最大化小睡效果。小睡应出现在警觉性低谷,通常在自然醒来后 7-8 小时。小睡新手常错过正确的时机!以 7 小时为起点,万一你因故迟到,还能留出 60 分钟的余量。
- 只在小睡之后喝咖啡或其他含咖啡因饮料。系统中哪怕极少量的咖啡因都可能干扰睡眠质量。
- 如果你在小睡前饮酒,它应在你入睡时基本被代谢(即离开你的系统)。
- 如果你小睡超过 100 分钟,你可能需要在夜间睡更多(或你小睡得太晚)。
- 小睡前 1-2 小时避免压力。即便你喜欢的事也可能让你兴奋,使你更难获得小睡的好处。
- 运动有益。尽量在小睡前至少 45 分钟完成运动。如果你没能冷静下来,小睡可能过早结束。
- 推荐小睡前进餐。除非你的医生另有说法,你一天的主餐实际上应在小睡前(在清醒日第 6 小时左右)!
- 推荐小睡前的性爱。
- 坚持你的仪式(例如坚持你最佳的顺序:运动、啤酒、洗澡、进餐、安静的地方、小睡、音乐等)。
- 如果上述建议不起作用,你可能需要一个月左右的尝试。精神减慢在这里至关重要!许多人直到某些情况把他们带入这个习惯(例如心脏状况诊断)才发现小睡的好处。即便你无法入睡,你可能仍需要小睡!无法入睡可能只是习惯或你无法忘记一天担忧的结果。你可能根本没有完全理解自己实际的睡眠需求。
- 如果一个月尝试后这一切仍不能让你在 10 分钟内入睡,你大概可以安全地永久放弃小睡。
小睡迷思(Napping myths)
迷思 1:只有懒人才小睡(Myth #1: Only lazy people take naps)
在一些文化中,这个有害的迷思让人觉得需要小睡是软弱的羞耻。这个迷思必须立即废除。小睡对认知功能和生产力有巨大影响。如果你想小睡,就小睡并自豪地宣布。你在做一件聪明的事。小睡让你更聪明!
当纽特·金里奇(Newt Gingrich)在镜头前被抓到睡着时,评论者把手指向所有错误的方向:他老了、他对竞选疲倦了、他可能患阿尔茨海默病等。金里奇原定通过卫星连线向美国以色列公共事务委员会直播。在 Panetta 之前的演讲期间,他选择坐姿快速小睡。他的年龄或健康在这里不需要扮演任何角色。事实上,纽特在做一件聪明的事:在出现在要求高的观众面前之前清理大脑。然而,尽管可用时间很短,他还是设法进入了深睡眠,醒来时有明显的睡眠惰性迹象,不知道自己在哪、要做什么!他恢复得相当快,用攻击激进伊斯兰的套话。那些睡眠惰性症状不需要表明他对竞选疲倦,或他的睡眠剥夺是许多天睡眠债的延续。哪怕一晚没睡也足以把任何人置于他的境地。他作为政治老手反而对他不利。没有新手能克服公开睡觉的压力来获得几分钟睡眠。金里奇唯一明显的错误是前一晚没有获得完整的睡眠。更多内容见:Why naps cause sleep inertia?
迷思 2:小睡就是小睡就是小睡(Myth #2: A nap is a nap is a nap)
这个迷思说每一次小睡都是好小睡,何时进行、持续多久都无所谓。这个迷思深植于经验不足的小睡者的心理,他们常常没有意识到在约 24 小时周期中循环穿过人体的无数基因、代谢、神经和激素过程。在”最佳小睡时机”一节中,我对昼夜节律周期进行了划分,并对每个选定周期点进行小睡时发生的过程作了简短分析。在不同昼夜节律周期点的小睡,天差地别。有的令人神清气爽,有的是浪费时间,有的可能不健康(或至少低效),有的会持续数小时!
迷思 3:一定要从第 2 阶段 NREM 中醒来(Myth #3: Make sure you wake up from Stage 2 NREM)
一些小睡”专家”会告诉你用闹钟确保在 15-20 分钟后从第 2 阶段 NREM 醒来。据说,更长的小睡会让你进入深睡眠(第 3/4 阶段 NREM),醒来会昏沉。实际上,决定小睡时长和效果的是小睡相对于昼夜节律周期的时机,以及先前的睡眠剥夺和 REM 睡眠亏欠。在某些情况下,用闹钟切断小睡可能有些令人神清气爽,并防止时机不当的延长小睡的涟漪。然而,选择恰当的小睡时间总是更好的,通常在主观日的第 7-8 小时。这转化为自由睡眠中醒来后 7-8 小时。然而,在睡眠剥夺或睡眠周期错位的条件下,较早小睡甚至完全跳过小睡以帮助周期重新同步更安全。
你听过一个故事,据说爱因斯坦夹着铅笔小睡,以便铅笔一掉就醒来?我怀疑一个伟大的天才会在规律基础上犯这个错误。我相信他有机会比较时机恰当的自然小睡与被打断小睡的价值。也许铅笔确实掉了。一次?也许天才的大脑因此被剥夺了某些新的创造性洞见?或者反过来,他因被打断的思绪而受到启发?也许那只是喝茶时讲的好故事?无论真相如何,不要效仿这个例子!让你的大脑决定小睡应该持续多久!
迷思 4:昼夜节律周期可以被忽略或废除(Myth #4: The circadian cycle can be ignored or abolished)
一个有害的迷思说,我们可以忽略昼夜节律周期,使睡眠简化为一维的内稳态过程。这个迷思源于对睡眠双过程本质的缺乏理解。它使许多人相信多相睡眠是良好的长期生活方式选择。这个迷思源于对主要昼夜节律睡眠组分压倒性力量的缺乏认识。因此,这个迷思带有一种信念,即小睡可以在任何与上次小睡足够远的任意时间被随意引发。
看看下面这张借助 SuperMemo 获得的惊人图片:
该图显示了人类昼夜节律周期强大的双相本性。横轴显示昼夜节律时间,即从相位 0(预测的”夜晚结束”时间)流逝的时间。预测来自 SleepChart 采用的昼夜节律模型,并源自 SuperMemo 中收集的睡眠日志。红线是使用为 SuperMemo 睡眠优化而开发的双过程睡眠模型(该模型受 Alexander A. Borbély 和 Peter Achermann 类似工作的启发)从同一睡眠日志数据推导的预测警觉性。警觉性是两个睡眠驱力过程状态的合成:
- 内稳态过程,以及
- 昼夜节律过程。
蓝点是从 SuperMemo 实际学习过程中获取的回忆数据。换言之:红色是模型,蓝色是数据。两者讲述同样的故事!对于不相信科学模型的怀疑者,不带偏见的蓝点数据应是决定性的论据。该图明确表示,我们一天中有两个主要的警觉性峰值。它也清楚地指出,只有两个适合睡眠和小睡的低谷。
最佳小睡时机(Best nap timing)
小睡是疲倦大脑的恩赐。然而,如果在错误的时间进行,也可能搞乱你的睡眠周期。许多人认为小睡就是小睡就是小睡,无论时机如何都会让头脑神清气爽。这是错误的。理解小睡的最佳昼夜节律时机,对小睡成为你的朋友而非敌人至关重要!对小睡普适性的信念激发了一个危险的 ideavirus:基于多相睡眠的生活方式。
小睡是一门技能。许多人即便困倦也无法小睡。测量自然醒来与小睡之间的时间,应有助于你优化小睡质量。最佳情况下,你的疲倦可能甚至不明显到足以轻松猜出最佳时机。如果你测量夜间睡眠与小睡之间的时间,你会注意到长度总是一样的(微小变化取决于夜间睡眠质量)。换言之,测量帮助你在中午不感到疲倦的日子也能计算出昼夜节律低谷的时机。你可能会想,那干嘛还要小睡?认知能力的提升值得投资(好日子可能只需 10-20 分钟)。
在健康的双相睡眠中,午睡时段的小睡是精神能量和创造力的绝佳提升。重要的是要知道,小睡时机不应由墙上挂的时钟决定。你的小睡应在自然醒来时间后约 7-8 小时。精确地说,只有你能通过比较早一点或晚一点小睡来确定这个值。如果你用闹钟缩短睡眠,或比平时更早入睡(例如因为疲惫的一天),或将入睡推迟到自然睡眠时刻之后,这个最佳值可能不成立。
小睡的昼夜节律时机(Circadian timing of naps)
为了优化小睡,你需要理解睡眠周期相位的影响。下面我解释在不同相位小睡会发生什么。在下文中,相位 7 小睡指在自然小时的自然睡眠自然醒来后 7 小时进行的小睡。请参考下图,它说明了人类睡眠倾向的双相本性。该图显示只有一个适合睡眠的最佳警觉性低谷(红色),通常在第 7-8 小时:
相位 0:醒来时间(Phase 0: Waking time)
在相位 0 小睡是在醒来后立即进行的小睡,即上图中的昼夜节律时间 0。在相位 0 小睡是可能的,很大程度上取决于先前睡眠的历史。正常夜间睡眠后的相位 0 小睡,可以被视为夜间睡眠的补充(如果夜间睡眠不够有效)。这种小睡在睡眠模型中与夜间睡眠合并,是在夜间睡眠被打断(例如噪音、膀胱满、健康问题等)时延长夜间睡眠的有效方式。不眠之夜后的相位 0 小睡可作为夜间睡眠的低效替代。这种睡眠会短、不令人神清气爽并留下睡眠债,还会在昼夜节律系统中引入不受欢迎的振荡,可能需要几天才能清除。这种睡眠常被夜班工作者用来获得一天的精神提升。它仍远比完全不睡好。规则很简单:如果你在相位 0 困倦,就随意小睡。你的大脑显然需要更多睡眠。
相位 3:创造时间(Phase 3: Creativity time)
在相位 3 小睡在健康、调节良好的系统中不应可能(见上图横轴第 3 小时的红色峰值)。这个时间成功睡眠是睡眠剥夺、低质量睡眠(例如因睡眠呼吸暂停)、错误相位睡眠(例如过早入睡)、睡眠障碍(例如发作性睡病)等的迹象。这可能是所有时段中最难小睡的。然而,我不知道这种小睡对健康或睡眠控制系统有什么不良影响。
相位 5:午睡前(Phase 5: Pre-siesta)
在午睡前时段小睡是可能的。然而,这种小睡可能短,且不如相位 7 小睡令人神清气爽。由于昼夜节律原因,它们也更可能富含 REM。这些早小睡可能推荐给患有入睡性失眠、但仍想在警觉性和创造力方面提升下半天的人。这些小睡也可以”匆忙”执行,因为它们持续时间短,适用于不希望较长小睡或较晚时机不适合当天日程的情况。
相位 7:午睡(Phase 7: Siesta)
小睡的完美时间。如图所示,这是脑力表现处于中午低谷的时段(瞄准相位 7 以确保即便迟到,小睡仍落在低谷内)。低谷由丰盛的午餐引起的说法是错误的(尽管餐食对睡眠控制有很大影响)。低谷是昼夜节律波的自然表达。这个昼夜节律低谷时段大致与主观夜晚低谷在约 12 小时的偏移处同一时钟时间(例如,如果你夜晚的中点落在凌晨 3 点,下午 3 点的小睡可能最有效)。这在 “How to nap” 中有很好的解释。午睡的好处已被大量研究证实,并在世界许多地区实践了几个世纪。随着人类生产力日益依赖我们的创造力,它肯定会渗透到企业界。
相位 11:傍晚(Phase 11: Evening)
这不是小睡的好时间。在健康的周期中,小睡可能难以实现或不可能。然而,即便轻微程度的睡眠剥夺也会产生一次可能触发负责整夜睡眠的控制机制的小睡。晚小睡可能富含 NREM 睡眠,并剥夺你夜间睡眠中至关重要的 SWS 组分(Werth 等,199652)。这些小睡可以比午睡长得多,会让你昏沉。最糟的是,它们会加剧失眠。不幸的是,这是很大一部分学生采取的小睡类型!被迫在不成体统的清晨早起上学,孩子和学生半梦半醒地挣扎着度过上课时间,学习进展微乎其微。在这种状态下学习只会放大对学校相当普遍的憎恨。相位 11 小睡于是成了熬过一天、在傍晚进行一些实际学习的唯一方式。生物钟把主观夜晚移到早晨时段。正面的副作用是傍晚可以充满有效的学习。负面的副作用是学生发现不可能在凌晨 3-4 点前入睡,并在主观夜晚中段响起的闹钟声中迎接新的明亮上学日。这延续了痛苦和学校憎恨的循环。没有人估计过这一现象的全球后果,包括对众所周知问题缠身的青少年态度的影响。也没有人提出实际的解决方案(推迟上课时间通常导致孩子们通过同时推迟就寝时间来”适应”新周期)。我在这里也无法推荐解决方案。跳过傍晚小睡可能对夜间睡眠质量和昼夜节律周期更早相位的稳定更好,然而那将有效地剥夺这些学生唯一能学习的时间。这些傍晚小睡也是这些年轻大脑渴望的自由睡眠的唯一微薄替代,是大脑唯一获得所需的时间。如果我要回答:小睡还是不小睡,我可能不得不承认在大多数情况下傍晚小睡是较小的恶。
相位 13:睡前(Phase 13: Pre-sleep)
这是小睡特别糟糕的时间。在这个时间启动小睡应该相对容易。然而,睡前小睡可能产生以下不受欢迎的结果之一:长小睡-短夜间或长夜间-早醒(取决于睡眠控制系统的当前状态)。睡前小睡可能触发夜间睡眠序列。然而,这个序列并非不可打破,可能导致早醒加上重新启动睡眠的困难。如果内稳态睡眠过程产生大量困倦,而昼夜节律过程尚未为夜间睡眠成熟,这尤其可能。结果,这种睡前小睡产生的总睡眠可能少于正常夜间睡眠。这种长小睡-短夜间不能完全履行睡眠的生理功能。因此,你第二天的警觉性水平可能大幅下降。睡前小睡较不不幸的结果是,如果你成功触发不间断的夜间睡眠序列。然而,你可能在昼夜节律睡眠功能完成之前过早耗尽内稳态过程,可能比平时更早醒来。这是长夜间-早醒的结果,考虑到过早的睡眠常常比普通夜间睡眠长得多,这些夜晚惊人地不令人神清气爽。这种低睡眠效率的原因可能是 REM 睡眠的稀缺,而 REM 强烈依赖昼夜节律。此外,由于昼夜节律原因,你的早晨可能异常困倦!
相位 15:分段睡眠(Phase 15: Segmented sleep)
相位 15 小睡应被视为”早夜间睡眠”。如果你在这个时间入睡,可以预期以下任何一种(取决于睡眠债程度):
- 如果你没有睡眠债:床上一些无效时间,因为你可能无法入睡。
- 如相位 13 睡眠的睡前结果:长小睡加短夜间,或长夜间加早醒。这个结果可能让你更不神清气爽。
- 分段睡眠:你会入睡,但在夜里醒来 1-2 小时,然后重新入睡。这个幸运的结果会让你相当神清气爽。
- 长夜间:在罕见情况下,尤其是如果你睡眠剥夺,早的就寝时间会导致美好、长、令人神清气爽的夜间睡眠。
由于相位 15 睡眠的不确定性,它应只在睡眠剥夺条件下使用(这提供了良好结果的较高机会)。否则,早的就寝时间在最好情况下可能无效,在最坏情况下对睡眠质量有害。
相位 18-24:夜间睡眠(Phase 18-24: Night sleep)
如果你试图在相位 18-24 小睡,你注定会触发正常的健康夜间睡眠。这没问题,只要你不是带着 20-40 分钟内停止过程的邪恶意图去”小睡”。这就是多相睡眠的痛苦最难承受的地方。正如 Stampi 博士二十年前注意到的,最令人沮丧的不是保持清醒或入睡的问题。多相生活最痛苦的部分是,当你的大脑想要触发夜间睡眠序列,而一位多相信徒固执地不允许它(Stampi,199253)!这与任何对”全有或全无”生理过程(排尿、排便、性高潮、吞咽、心跳、打喷嚏、咳嗽、分娩等)的突然中止一样糟糕。许多多相博主注意到:“我发现当我的小睡变长时,我就变得昏沉。所以我尽量把它们控制在 20 分钟以内”。废话!如果你不启动夜间睡眠序列,你就不会承受被打断的痛苦。那干嘛还要小睡呢?推迟排便比中途停止它更容易。我遇到的最不寻常的夜间小睡控制方法是……”我在床上放很多杂物。那让我的小睡保持简短”!
重要!
- 小睡相位指的是最佳自然醒来时间,而非实际醒来时间! 例如,在过早的睡眠或被打断的睡眠中,高效小睡可以在醒来后 10 小时进行。如果你的睡眠不完全自然,你需要对内在昼夜节律周期(可能在后台独立于你实际睡眠时段运行)进行修正。不幸的是,绝大多数有睡眠相位问题的人不知道如何高效测量和绘制自己的自然周期。
- 两个警觉性低谷在生物学上不同! 如后所示,只有夜间低谷能产生典型的、漫长的周期性 NREM-REM 交互,并伴随 REM 比例的逐渐增加。主观夜晚时段以褪黑素释放的特征性增加为标志。如双相睡眠图所示,午睡时长比自然夜间睡眠短 4-20 倍。相位响应由先于或紧随夜间睡眠的刺激引发。然而,同样的刺激可能影响午睡小睡的时机,而午睡反过来可能对周期相位有间接影响。
- 小睡时机将决定其结构。 一些研究者认为,由于小睡时机决定了 NREM 与 REM 的比例,这也会影响小睡对特定心理任务的效率。如果你想为特定任务最大化价值/时间,这可能是真的。然而,在睡眠优化中,你应始终追求最大总价值。这就是为什么最好的小睡是与中午昼夜节律警觉性低谷良好对齐的小睡。
小睡的良好条件很重要。在半卧位、嘈杂房间或明亮灯光下的小睡,也会带来警觉性好处(前提是你真的能入睡,也许通过第 1 阶段 NREM)。然而,在有利于睡眠的环境中的小睡通常更长,也远更令人神清气爽。
一天一次小睡足够(One nap per day is enough)
许多人相信,每一次额外的成功小睡都对恢复精神能量珍贵地有帮助。在睡眠不剥夺的正常睡眠者中,额外的小睡确实可能带来增加的警觉性和改善的脑力表现。然而,在健康的作息下,午睡时段以外的所有小睡都应非常难以实现。如果睡眠的目标被定义为实现最大创造性生产力,且夜间睡眠能健康地走完全程(即没有睡眠剥夺),那么在午睡时段以外任何时间的小睡尝试都是浪费。这是因为入睡应该困难,仅仅闭眼休息产生不了一次成功小睡神经收益的一小部分。此外,即便成功,早上强行插入的额外小睡可能干扰下午小睡。同样,傍晚小睡可能缩短夜间睡眠。这些额外小睡可能带来表现的渐进改善,但会降低睡眠的整体效率,并可能在昼夜节律系统中引起涟漪。我们的双相本性明确表明,我们应致力于下午(清醒第 7 小时)的单一小睡。对一些人,即便这也太多,单相模式是他们的最佳。
许多年轻的创造型个人提出自己的设计师睡眠作息。我常收到提交新睡眠模式建议的邮件。例如,三相睡眠:一次 6 小时的主睡眠(00:00-06:00),加上餐后两次 30 分钟小睡(12:00-12:30 和 18:00-18:30)。与大多数让睡眠系统按设计运作的人为方式一样,这种作息不太可能高效。大多数人是强烈双相的,只有双相或单相睡眠对成年人有效。然而,如果扔掉第二次小睡,这个建议就相当接近自然的双相节律:0:00-6:00 和 13:00-14:00。即便如此,设计师作息应始终避免。唯一的例外是那些近似 SleepChart 在自由睡眠中显示的设计。由于人们在睡眠控制系统各种参数上不同,那些非常规律的睡眠者确实可以考虑把特定时机写进作息,只要时机来自他们实际睡眠模式的测量。如果设计师作息中的睡眠事件未与昼夜节律需求对齐,它们常常导致昼夜节律混乱。
如果你想睡好且高效,选择双相、单相或自由睡眠中对你最有效、你能负担的。与日光周期同步的自由睡眠是最健康的,将带来最高生产力。一旦你自由运行,你将迅速确定自己偏好双相还是单相睡眠。
多相睡眠(Polyphasic sleep)
加速回报定律(The law of accelerating returns)
我们生活在加速的加速时代。摩尔定律让世界更小、更快、更互联、更高效。我们现在能够触摸和感受 Kurzweil 的概括:加速回报定律。快节奏的年轻一代渴望更多:更多乐趣、更多信息、更多成就、更多教育,以及……更多清醒时间。看到这么多人为了缩短睡眠、提高生产力而竭尽全力,相当令人惊叹。年轻的自我实验者不断用闹钟缩短睡眠、使用受控物质、规律通宵,或试图把睡眠时间用于”有用目的”(例如睡眠中学习)。
在一位试图通过少睡来拯救生命的年轻企业家的博客中,我读到(粗体强调是我的):“睡眠不是我的朋友。作为一个初露头角的年轻企业家,我渴望用更少睡眠、更多清醒时刻来生活。我总觉得床上的那些时刻本可用于更崇高的目的。[……] 任何优秀的工业技术专家都知道,要控制某事物就必须测量它。所以我写下一年来每天入睡和醒来的时间。测量给了我一个改进的基准”。随后是一份值得称赞的、一年的测量记录,产生了一张健康规律睡眠的漂亮图表,平均每晚令人羡慕的 8 小时(夏季略少)。睡眠专家可能会惊呼:“对你好!继续保持!” 但这位初露头角的企业家继续”改进”数百万年的演化。“这总共是 112 天睡眠和 253 天清醒。换言之,我睡掉了这一年的 31%,[……] 我做了一些实验,看看能否减少睡眠时间。例如,我曾严格地把每晚睡眠限制在 5 小时……持续了大约 2 周,我让自己发烧了。然后我尝试每周通宵一次,能坚持几周就几周。那也没持续多久。主要是因为通宵后的第二天总是如此低效,以至于我多醒着的时间并没有带来净生产力增长。[……] 我还没有找到一种持续少睡的好方法。” 我纳闷为什么一个优秀的工业技术专家不愿花 5 分钟谷歌一下睡眠的功能和崇高目的。毕竟,你不需要生物学荣誉学位就能知道,如果身体做某件事,几乎总是为了一个重要目的。
除了对更多生产力和更多清醒时间的渴望,人类心智的造迷力量如今被全能的互联网怪诞地放大。如果一个想法能让生活更好或更可忍受,它一旦被发明就会迅速拥有自己的互联网生命。按照模因学(memetic science)的规则,这个想法生长、突变并演化。它自由地汲取科学以及谣言、自我实验,以及为利益愿以真相换利润的不择手段的偏见来源。它像滚雪球一样,在它碾过毫无准备的心智时,添加新的、令人愉悦的事实和假设。像一个新的救世主,它拖着新的追随者、倡导者、使徒和死忠游击队员,准备为事业的最终胜利贡献力量。
大约 2000 年,一个新的模因(meme)在网上的几个博客中冒出:Uberman 睡眠作息(The Uberman’s Sleep Schedule)。由于我对睡眠在记忆和学习中作用的兴趣,这个模因没过多久就到达我的收件箱。随着这个概念每月膨胀,我别无选择只能表态。
Uberman 睡眠作息(The Uberman’s Sleep Schedule)
Uberman 睡眠作息背后的想法是,通过在一天中分 6 等份总共只睡 3 小时来获得清醒时间。该方案有许多变体,由尝试按此作息睡眠的人提出。该作息本应压缩生理上不那么重要的睡眠阶段,并 homeostatically 上调对心理健康至关重要的阶段。Uberman 睡眠作息由一位躲在昵称 PureDoxyk 后的女性在这篇 Everything2 博客中提出。该博客报道了一次睡眠实验,坦率承认 Uberman 作息与实验者的作息和目标不兼容。然而这个模因在 2002 年的一篇 Kuro5hin 文章中被拾起。用简单且结构良好的语言表达,这次它被注意到了。同样,该文以*“Uberman 睡眠作息是一种潜在危险的增加清醒时间的方式”*结尾。这并未阻止一波新的追随者准备好获得多年的清醒时间。该概念的吸引人主题是,确实,如果你成功每天睡 3 小时而非规定的 8 小时,从 20 岁开始,你在平均西方寿命中将获得超过 11 年。这个想法非常有吸引力。难怪它似乎在相当长一段时间内持续获得动力。
多相睡眠(Polyphasic sleep)
越来越频繁地,Uberman 睡眠作息被称为**多相睡眠(polyphasic sleep)**(该术语由意大利时间生物学家 Claudio Stampi 博士的研究和著作推广)。睡眠研究者把多相睡眠视为一种睡眠模式的变体,与单相或双相睡眠相对。在单相睡眠中,个体或动物在 24 小时的单一清醒-睡眠周期中以单一区块睡眠。在自然的双相睡眠中,24 小时内有两个睡眠区块:夜间睡眠和典型的拉丁午睡(“第 7 小时小睡”)。
多相睡眠在动物界相当普遍。在系统发生的重演中,人类婴儿也多相睡眠,并逐渐失去其小睡倾向,直到约一岁时大致成为双相。人类成年人,与所有类人猿一样,主要是双相的。尽管大多数西方人不规律小睡,他们的警觉性在主观日中段显示低谷。这个低谷在自由睡眠条件下可合并成短暂的睡眠事件。
Uberman 睡眠作息背后的理论是,通过一些努力,我们可以训练大脑沿古老的多相周期睡眠,并在过程中获得大量清醒时间,主要通过抛弃不那么重要的睡眠阶段(例如缩短 NREM 第 1 阶段,它似乎只是通往更”有用”的慢波睡眠阶段的过渡状态)。这一理论有缺陷,因为它没有考虑主观夜晚昼夜节律顶点的量级。
是否应该多相睡眠(To sleep or not to sleep polyphasically)
在介绍了热情倡导者眼中的多相睡眠后,让我们看看其生理根源和影响。随着每个月过去,我们积累了关于睡眠在记忆和创造力中关键作用的大量证据。此外,我们大多数人对”没有睡眠就几乎没有智力成就的机会”有良好理解。更重要的是,我们发现不借助外力保持清醒超过 2 天很困难。尽管有文献记载的超人成就,如 Peter Tripp(1959)、Randy Gardner(1965)和 Tony Wright(2007)分别(半)清醒 8、11 和 11 天,大多数凡人甚至无法熬过前 48 小时的清醒,不可避免地沦为睡意的猎物。
脑电图(EEG)和体动记录仪(actigraphy)测量表明,人类基本上是双相的。在主观夜晚有单一的、强大的睡眠驱力,在主观日中段有单一的警觉性低谷。脑电图测量得到许多其他生理变量的证实,如体温测量、血液中皮质醇水平、唾液中褪黑素水平、其他激素水平、血压、基因转录、免疫细胞活动、主观警觉性,以及无数其他参数。2007 年,我终于能在 SuperMemo 中看到记忆回忆和巩固的昼夜节律变化的同样效应。
人类 24 小时周期性的根源在于大脑中视交叉上核(SCN)的活动,它由运行经典反馈回路的 24 小时基因转录变化周期驱动。负责昼夜节律周期性的基因中的微小突变可能延长或缩短昼夜节律周期,也可能导致完全的无节律性。许多这类突变已在果蝇和小鼠中被研究。导致睡眠相位障碍的人类突变也已知晓(如家族性 ASPS)。然而,这些突变罕见,对绝大多数健康人,周期长度略长于 24 小时。Charles Czeisler 博士在所研究的条件和样本内,测量其为 24.2 小时,个体间变化惊人地小(Czeisler 等,199954)。昼夜节律周期(包括基因转录和 SCN 活动)可以每天被轻微推动和移动,移动程度由相位响应曲线(PRC)决定,并需要相位移动刺激的非常精确的时机(Khalsa 等,200355)。换言之,通过光、体力活动或社交互动等刺激,我们可以轻微移动最大困倦感的周期。尽管 PRC 的精确测量提到,单一强刺激一天可移动多达 3 小时,但在实践中,很难每天移动超过 1 小时。我们都试图适应 24 小时日时每天得到一点向后的推力。这种每日重置对应用睡眠卫生原则的人来说是无痛的,它在早晨随光、活动和/或压力发生。越来越多的人口使用闹钟来完成本应由阳光和活动自然完成的工作。这不是健康的解决方案,通常由我们电照明的生活方式(晚间电视、晚间阅读、晚间互联网、晚间聚会等)所迫。对于那些失相位的人,把睡眠作息推到更晚时段比推回去更容易。这种不对称源于我们可以有意识地控制清醒时段,而这只能用于向前移动。让自己熬夜很容易,让自己早起则困难得多。自然,闹钟可以用于后者,但在时间疗法和健康睡眠中应避免使用闹钟,因为闹钟对睡眠周期进展的破坏性影响。
虽然可以移动睡眠相位,但我们不知道任何生物机制可以在不引起一定程度睡眠剥夺的情况下显著缩短健康睡眠区块的长度。最短的有效睡眠可以在自由睡眠中通过强烈的清晨重置刺激和强烈的傍晚相位延迟刺激实现。然而,即便自然刺激也可能引起一定程度的睡眠剥夺。移动睡眠相位对主睡眠区块长度的影响相对较小,变化与移动程度成正比,符号相同(即移动延迟减少主观夜间睡眠长度)。最重要的是,变化在移动后不久就回到基线。这说明了睡眠控制机制的内稳态本质,只要移动足够小且由时机恰当的相位移动刺激产生,它就会在很短时间内通过在新偏移处稳定新的睡眠作息来响应相位移动刺激。对睡眠模式更果断的干预可能导致昼夜节律混乱。在西伯利亚仓鼠中,它甚至可能引起无节律性(Ruby 等,199656)。这些剧烈变化可能有严重的健康影响,且可能难以缓慢逆转。
影响睡眠模式的自然刺激的明确效应导致一个即时结论:声称人类可以适应任何睡眠模式是错误的。作息的突然改变,如轮班工作,可能导致睡眠控制机制的灾难性破坏。北美 25% 的人口可能以轮班作息的变体工作。许多轮班工作者从未把睡眠模式适应轮班模式,他们有时在生物钟有害脱节的情况下部分工作,一旦轮班回到他们偏好的时段就回到令人神清气爽的睡眠。在最坏的情况下,工作时间的持续移动导致各生理变量之间失去同步,工作者从未获得任何高质量睡眠。这把个人推上一条直接通往一连串健康问题的道路,包括心脏疾病、免疫系统抑制、糖尿病、胃肠道疾病、肥胖、抑郁、慢性疲劳、睡眠障碍等。轮班工作者也面临更高的事故和家庭问题风险(如更高的离婚率)。轮班工作设计应应用时间生物学定律,以最小化轮班工作对健康的不利影响。通常,让工人持续上夜班比通过无休止的作息轮换周期每周诱发规律的睡眠破坏和压力要好。
看来,多相睡眠遇到的问题与时差或轮班工作中看到的完全相同。成年人的生物钟不适应单相或双相以外的睡眠模式。换言之,唯一已知的健康替代方案是:
- 夜间单一 6-8 小时的睡眠区块,或
- 5-7 小时夜间睡眠加上 15-90 分钟午睡小睡。这些数字在整个人口中差异很大,没有适合每个人的单一推荐睡眠剂量。
如果对生物 clock 施加一定程度的压力(例如比身体最佳时间更晚入睡),中午小睡可以作为代偿缓冲,抵消睡眠剥夺。在这种条件下,小睡可能比通常的 15-30 分钟更长。对夜间睡眠施加的压力越大,午睡越长。类似的双相整合也可以在大鼠中实验性产生。看来,在足够压力下,小睡可能变得比夜间睡眠更长,有效地把睡眠模式反转 12 小时。这一效应证实了人类睡眠双相本性的一个重要方面,而这一点目前的睡眠模型尚未完全考虑。在罕见情况下,个体可能学会分两个 3-4 小时的区块睡眠。然而,在绝大多数情况下,睡眠在 24 小时内以两个相等区块出现的模式是不稳定的。换言之,按比例双相作息的个体很快回到长夜间短午睡,或回到单相睡眠。通常,黑暗期间发生的睡眠部分扮演夜间睡眠的角色。然而,更可能的是,这个角色由在建立按比例双相模式之前更长的那部分睡眠承担。这再次表明,双相模式中两个睡眠区块之间潜在的生理不对称。换言之,身体记得哪个睡眠区块是主观夜晚区块,即便那个区块碰巧发生在日间。
通过睡眠剥夺,利用困倦的内稳态组分,多相睡眠者可以增加白天的小睡次数。然而,一次夜间睡眠加多次白天小睡的模式高度不稳定,只能在永无止境的睡眠剥夺下维持。自然,如果你碰巧使用闹钟,你可以轻易地在主观夜晚的昼夜节律低谷期间运行多次”小睡”。这在主观白天是不可能的(除非在极端睡眠剥夺条件下)。在某种程度上,闹钟也可以被你的内部闹钟替代(例如想”我必须在 20 分钟内起床”)。在类似条件下执行的”小睡”都不能完成自然睡眠的工作,它们不仅很大程度上是浪费时间,还有助于拆解你的睡眠控制机制。Stampi 博士关于多相睡眠的研究也明确指出了睡眠的禁区,在那里没有大量睡眠剥夺很难启动小睡。这些禁区在双相节律上映射良好,主观傍晚先于核心夜间睡眠的小睡对休息良好的个体尤其无效。所有上述发现不可避免地导致一个结论:不可能在保持多相睡眠作息的同时保持高警觉性和/或创造力!如后所示,实践对多相睡眠对创造型个体的不可行性同样不留情面。
轶事证据似乎表明,高度创造性的个体在双相睡眠模式下表现最佳。然而,最大化创造力和警觉性的唯一有效经验法则是:只有当你感到困倦时才睡。当这个规则被应用时,个体可能落入多种不同的作息。他们在任何这些典型的单相和双相模式中可能都相当有效:典型的 7+2 或 6+1、长睡眠者的 9+0、短睡眠者的 4+1,甚至 4+0 等。只有你能确定哪种作息对你最佳。然而,你可以预期,如果你是正常的健康个体,这种作息不会是多相(双相以外)。如果你尝试 3+0.5+0.5+0.5,你要么严重睡眠剥夺(即只能靠闹钟维持作息),要么会回到 3+0.5,或更可能地,回到标准的 6+1 模式。在不睡眠剥夺的情况下把小睡挂接到超日节律(ultradian rhythm)的可能性是个迷思。
Uberman 大爆炸五年后(5 years since the Uberman Big Bang)
在 2002-2005 年,我注意到对 Uberman 睡眠作息兴趣的指数级增长。我不断收到越来越多询问 Uberman 对健康和学习影响的邮件。因此,我写了《Polyphasic Sleep: Facts and Myths》。在随后的五年里,我收到了约 500 封邮件,并亲自与许多多相睡眠信徒接触。在我有机会监测的尝试中,全部不成功。原文的一些批评者声称他们确实多相睡眠,但我没有收到可作为验证基础的数据。从大量邮件中得出的最有趣结论是,人们转向多相睡眠,更多是因为先前的睡眠问题,而非对成就的渴望。他们常把多相睡眠视为所有睡眠问题的万灵药,这种认知被多个博客声明放大。我收到了几份 SleepChart 数据提交,展示了与多相睡眠斗争有多困难。诚然,我对一些实验者展现的决心印象深刻。随着最新版 SleepChart 能建模睡眠倾向的变化,它提供了对多相睡眠中发生的相位移动混乱的一些洞见。随着每一天过去,我们对多相睡眠及其对健康潜在影响的了解越来越多。对多相睡眠作为生活方式的支持者来说,消息并不好。
我收到的关于多相睡眠文章的邮件大多是批评性的,但不应以此作为成功的衡量标准。重要的是有多少读者会同意我,而是有多少人因此放弃了多相睡眠的想法。在 500 封邮件中,我粗略估计其性质分布如下:50% 批评,40% 请求帮助实施多相睡眠,10% 感谢我警告不要采纳多相睡眠。10% 的转化率似乎很低,然而这相当于数百小时某人自己的时间,我确信它也转化为切实的健康收益。例如,大量多相尝试以感冒或流感告终,这必须反映这种睡眠作息对免疫系统的影响。Aaron 写的相当代表 10% 群体:“全天分散小睡的想法吸引了我,因为我一直遭受我无法恰当量化、但现在知道是 DSPS 的困扰。如果我不使用闹钟,并在疲倦时入睡,我看到我的睡眠/醒来时间每天显著移到更晚时段(晚几小时)。这对我一直是挫折的来源,我考虑用多相作息来帮助纠正问题。然而,在阅读《Polyphasic Sleep: Facts and Myths》后,我决定这将是真诚地浪费我的时间。” 对我文章的批评通常绕过科学论证,引用声称在多相睡眠上成功的人的博客。例如 Kop 写道:“有许多人成功适应了。[……] 你只是忽略了引用他们,而你只引用了失败的人。我认为这对你的读者非常不公平和误导。我可能听起来像一张破唱片,但即便你相信每个声称成功的人都在撒谎,你也应该让你的读者做这个选择,你绝对不应该完全省略你个人不同意的所有信息”。
睡眠剥夺中睡眠阶段的压缩(Compression of sleep stages in sleep deprivation)
Uberman 睡眠作息的一个迷思是,它使进入 REM 睡眠并完全跳过非 REM 睡眠阶段成为可能。这个迷思源自另一个错误主张,即暗示深睡眠的非必要性。我将忽略这些主张,因为它们与实验室发现完全相悖。相反,让我们关注多相睡眠中压缩睡眠阶段的可能性的更合理主张。
确实,睡眠剥夺的人比正常睡眠者更快进入深睡眠。经过一段睡眠剥夺后,不那么重要的睡眠阶段被压缩,而核心 SWS 占主导。此外,REM 剥夺会导致恢复时的 REM 上调。最初,睡眠系统会努力赶上未完成的 SWS,然后才赶上未完成的 REM 睡眠(常常只在第二个恢复夜)。那么似乎确实,睡眠剥夺后我们睡得更高效。此外,睡眠的内稳态控制可能并不非常擅长检测真实的神经睡眠需求,这是可能的。如果你看看我们的哺乳动物亲戚,你可能会惊讶,长颈鹿可以靠 2 小时睡眠过得很好,而蝙蝠可能需要 20 小时。聪明且学习快的象需要比不那么聪明的大型猫科动物少 4 倍的睡眠。行为观察会迅速让我们得出结论,睡眠量与记忆获取和神经计算的量和复杂性不直接相关。
那么我们可以假设,睡眠控制可能采用只与神经优化需求松散相关的辅助生理参数。也有可能,演化考虑了夜间对早期人科动物不是非常有用的活动时间这一事实。睡眠控制机制可能因此吸引了一些额外的生理功能,即使睡眠持续时间长于记忆巩固和优化所需,这些功能也可能提高生存。因此,所有睡眠机制以悠闲节奏进行,加上许多原本不需要失去意识的附加功能的可能性。
如果上述思考正确,我们确实可能在有利环境下用更短时间执行同样的神经工作。然而,对神经优化与内稳态睡眠控制之间联系的真实本质所知甚少。我们目前的知识似乎仍坚定地表明,只有在自由睡眠中,我们才能最大化创造力与睡眠成本的比率。换言之,没有证据表明,通过玩弄睡眠剥夺,你可以增加创造力。唯一可能的例外是,通过推迟睡眠 30-60 分钟带来的微小剥夺程度。更长的延迟会影响警觉性,超过可能被视为”收益”的范围。可能的是,在自由睡眠和稍微延迟的睡眠相位这两个极端之间,(1) 睡眠压缩带来的时间收益与 (2) 加速的内稳态困倦之间的权衡,会在某处产生一个最佳点。自然,这种对睡眠周期的微小推动与使用闹钟、破碎作息以及 Uberman 睡眠实验者典型的永无止境的与昏沉作斗争毫无关系。此外,即便这种微小的假设性干预,也会不可避免地导致相位移动,可能有众多负面副作用,最明显的是无法在一个很大程度上与日光同步的社会中有效运作。驯化良好、平衡的自由睡眠仍是你获得最大认知表现的最佳选择。
在多相作息引起的睡眠剥夺中,由于睡眠阶段压缩,REM 睡眠会更快出现。然而,慢波睡眠才是内稳态上调的主要目标,强烈地由先前清醒的持续时间决定。由于 REM 睡眠与昼夜节律相位关联得多,它在睡眠中的比例实际上会下降,尤其是在主观傍晚时段启动的小睡中。你可能想研究 Alexander A. Borbély 和 Peter Achermann 的睡眠模型,它们很好地解释了这些过程的机制。实验室发现似乎表明,REM 的下降在连续的睡眠剥夺日子里逐渐恢复到基线,但这种恢复从不完整。换言之,与自由睡眠作息相比,你在多相作息上得到的 REM 睡眠更少。这种 REM 睡眠节食既是绝对的(以分钟计),也是相对的(与深睡眠 NREM 相比)。如果你从一个小睡中被闹钟叫醒,REM 剥夺的问题会变得更加明显。使用闹钟时,你统计上更频繁地击中 REM 睡眠,因为它的比例几乎总是随睡眠时间增加而增加。这就是为什么多相睡眠者常在醒来时记得梦境。那不是你获得更多 REM 的迹象,而是你在破坏 REM 睡眠的迹象。 通过使用非常短的睡眠区块,由于强烈的第 4 阶段 NREM 内稳态上调取代你能获得的任何 REM,你对 REM 的影响进一步加剧。
在多相睡眠中获得更多 REM 是迷思。你会得到更少的 REM,你的创造力会下降。
如果你 (1) 不与困倦对抗、(2) 自然从小睡中醒来,睡眠破坏的问题不会发生。然而,没有闹钟的帮助,不可能规律地适应一个预先计划的多相作息。这是因为健康个体唯一稳定的睡眠模式是单相或双相。多相睡眠模式在嗜睡症、发作性睡病(narcolepsy)和其他睡眠障碍的各种情况下可能是稳定和可持续的。当睡眠控制系统被破坏、内稳态睡眠组分过度工作时,频繁小睡可能发生并被推荐(例如在感染、化疗等期间)。毋庸多言,在这些情况下总睡眠增加,这正是多相信徒试图避免的。睡眠碎片化也可能因压力、社交生活、兴奋、过早入睡等而发生。这些干扰偶尔允许一天自然发生超过一次小睡的日子。如果你放弃闹钟,你就去除了使多相睡眠不健康的主要罪魁。然而,没有闹钟,决定睡眠作息的是你的身体,而非你预先计划的”合理化”作息图。那个作息不会类似任何 Uberman。
如果你的目标是获得大量含 REM 的小睡,你可能想知道,有两次以上含扎实 REM 睡眠的小睡是发作性睡病的诊断标准。
睡眠与创造力:少即是多(Sleep and creativity: Less is more)
多相睡眠对你的健康和创造力有害。然而,即便你想最大化清醒状态下的时间,假设你在清醒时间需要合理的警觉性来做任何事,它也可能不是你的最佳选择!只有在接近实质性睡眠剥夺时,多相作息在这方面才可能优于双相作息(见:PureDoxyk 定律)。
一些人,如消防员或急诊外科医生,可能为他人牺牲睡眠。需要做出牺牲的人数可以通过设计良好的轮班工作减少。其余大多数人口应为最佳健康和最佳清醒时段创造性表现而优化睡眠。多相睡眠绝对不是这类优化目标的答案。
这已经不是吉萨金字塔的时代,那时设计者的天才必须与 5 万名沦为纯粹繁重劳力的苦力配对。随着我们迈向**知识经济**,正是警觉和创造性的头脑为大多数项目的成功提供基础。一分钟的洞见可能值一个世纪的铲挖! 可能是单一的创造性 eureka 产生了 E=mc²。可能连爱因斯坦自己也无法回溯他大脑产生那个公式的确切时刻,他也无法为他人制定类似成就的可靠处方。人类创造力主要是概率游戏,但它只在沃土上繁衍。顶尖的头脑在顶尖的状态下,结合顶尖的睡眠,是未来更多这类洞见的最佳公式。多相睡眠是那个公式的对立面!
如果你浏览多相实验者的博客,你会看到他们一次又一次选择”吸引人的活动”只为保持清醒。为什么他们宁愿见人或慢跑,而不去投入一个有挑战性的脑力项目?
Tony Wright 在他挑战吉尼斯睡眠剥夺纪录(11 天不睡)的尝试中意识到,他什么都做得了,除了写作。10 天不睡后,他的大脑无法胜任写哪怕几个字的挑战。他得出结论:事实证明,在睡眠剥夺状态下写作是最难做的事,因此我决定不再写作,所以这将是我的最后一条记录。
为什么在睡眠剥夺状态下学习一门困难的学科会如此费力?由于运动或社交互动刺激大脑的胺类唤醒中心,它们是内稳态睡眠驱力的有效制衡。大脑使用最后的资源调动较少使用的皮层部分,以补偿海马体和其他中央记忆区域的过载。在创造力上,你可能已经脑死亡,但你仍能见人或慢跑。
学习是内稳态困倦的有力贡献者。学习的催眠力量是睡眠与记忆之间最可见的联系之一。如果你有入睡问题,没有什么比学习更好的天然催眠剂了!不仅是被动阅读,而是主动学习!我所知的最佳内稳态安眠药是增量阅读(incremental reading)。自然,你需要困倦的昼夜节律组分才能让这颗”药丸”起效。否则,学习(或增量阅读)矛盾地是你最好的”创造力丸”。
昼夜节律相位决定了学习产生创造性热情(睡眠后)的正向神经反馈,或困倦(睡眠前)的负向神经反馈。
然而,可能面临风险的不仅是警觉性、创造力和长期健康。可以想象,大脑中的睡眠控制中心会受多相实验影响。研究者注意到轮班工作或其他强制作息模式能使生物钟失常的案例。一些人推测,Peter Tripp 因其不眠挑战遭受长期后果。多相作息在睡眠剥夺方面不那么剧烈,但在昼夜节律破坏方面更剧烈。Stampi 博士让 Francesco Jost 经历每天 3 小时睡眠、持续 2 个月的饮食,没有可测量的不良影响。然而,审视其他神经病理生理过程,我们可能会担心,确实有可能杀死负责 SWS 开关、REM-on 开关、REM-off 开关等核团中的细胞。我们知道,忽视心理卫生、抑郁、过度细胞活动、谷氨酸、皮质醇、缺氧和其他神经压力会导致细胞丢失。我们知道可以用光刺激在西伯利亚仓鼠中解耦昼夜节律周期(Ruby 等,199656)。只要这一领域仍是灰色地带,摆弄自己的睡眠作息就等于拿自己长期有效控制睡眠-清醒周期的能力来掷骰子。这可能和节食没有太大不同:一旦你让食欲控制中心失效,你就注定终生与节食和体重反弹斗争。最近研究表明,垃圾饮食如何对控制食欲的脑中心造成胶质损伤(Szwartz 等,201257)。我打赌,垃圾睡眠导致睡眠-清醒有效控制丧失的可能性非常高。机制相同:当你让一个脑中心过度运转,你就有受伤风险。我们可以在十几个生理情境中看到同样的机制。一些多相信徒报告实验结束后睡眠方式不同了。其中一些报告可能暗示昼夜节律周期的扁平化,这是老年人睡眠控制的特征。结论如下:
违背睡眠-清醒周期的自然进程,你冒着永久损害自己产生健康、规律、驯化且令人神清气爽睡眠能力的风险。
为什么少即是多?因为通过给大脑它想要的那么多睡眠,你在清醒时段可以远更有创造力和生产力。不仅是远更多。在多相睡眠者中,创造力可能下降一个数量级。就像顶级运动表现。错误的用餐时机可能夺走 Usain Bolt 的奥运金牌。不要让自己在追求智力卓越的竞争中被边缘化!
婴儿中的多相睡眠(Polyphasic sleep in babies)
新生儿多相睡眠。生物钟基因在子宫内发育早期就开始循环。第一个昼夜节律也在子宫内开始显现,并被驯化到母亲的昼夜节律周期(如踢腿、呼吸、心率等)。然而,昼夜节律睡眠-清醒周期只在出生后发育。SCN 在出生后以非常快的速度生长。例如,它只含有成年血管加压素(vasopressin)表达神经元数量的 13%(Swaab D.F. 等,199058)。一个假设说,是视觉系统与 SCN 之间的连接只在出生后发育(Swaab 等,199459)。在早产婴儿病房进行的研究表明,适度的明暗周期加速昼夜节律的发育,而持续光照有相反效果,包括减缓儿童整体生长和发育(Mirmiran 等,200060)。甚至有报告称一个足月婴儿在研究期间未发育出昼夜节律周期,可能是因为它是夜间唯一在充分光照下喂养的婴儿(McMillen 等,1991)。新生儿 SCN 及其传入神经的不成熟,导致他们在出生后第一个月无法把周期驯化到日光。与此同时,对夜间睡眠的一些偏好可能与子宫内驯化的周期和/或出生后对母乳喂养和母亲周期(包括共同睡眠)的驯化有关。
一些多相睡眠支持者声称,婴儿睡眠是最自然的睡眠方式,婴儿因社会训练而早早失去它。事实恰恰相反。新生儿在睡眠模式中没有可辨别的昼夜节律偏好。这些模式在生命的头 1-3 个月迅速发育,与训练关系不大。典型的双相昼夜节律的发育是一个基因编程的生物学过程,在正常照明和正常社会环境下很大程度上不可避免。
婴儿多相睡眠。他们的昼夜节律睡眠周期在生命的头 1-3 个月自然发育,与”社会训练”无关。自然光、母乳喂养和共同睡眠有助于健康昼夜节律周期的发育。
除了传播”社会训练”迷思外,多相睡眠支持者还忽视了这样一个事实:婴儿睡眠的小时数远多于所谓的多相睡眠者(例如 10-16 小时而非期望的 3 小时)。健康个体不可能持续多相睡眠,也不可能睡 16 小时,除非处于严重睡眠剥夺状态。婴儿不使用闹钟控制睡眠时机(除了他们的饥饿闹钟)。看看新生儿第一个月多相睡眠的示例图,注意睡眠事件因各种内稳态因素的汇合而不规律地出现:

在健康婴儿中,控制入睡的两个主要内稳态是睡眠和喂养。毋庸多说,没有规律 Uberman 模式的迹象。如果存在超日节律周期,它们表达微弱,难以过滤掉。另一方面,可以看到一组慢慢出现的昼夜节律偏好,尤其是在睡眠事件合并后。在本例中,睡眠事件密度在晚上 22 点 - 早上 5 点时段更高(更多见下一节)。
最后但同样重要的是,多相睡眠倡导者,尽管流传着广泛的多相迷思,却在一阶上丢失 REM 睡眠。另一方面,婴儿可能把高达 65% 的睡眠花在 REM 上,没有它,他们的大脑皮层甚至无法正确发育(如睡眠剥夺小猫所证明(Stryker 等,200110))。
婴儿中的超日节律振荡(Ultradian oscillations in babies)
SleepChart 的昼夜节律图可用于寻找婴儿多相睡眠阶段的超日节律性。所给的图对应生命的头 7 周,未显示清晰的超日节律振荡,尽管 3 小时倍数间隔的峰值占所有峰值间隔的 75%:
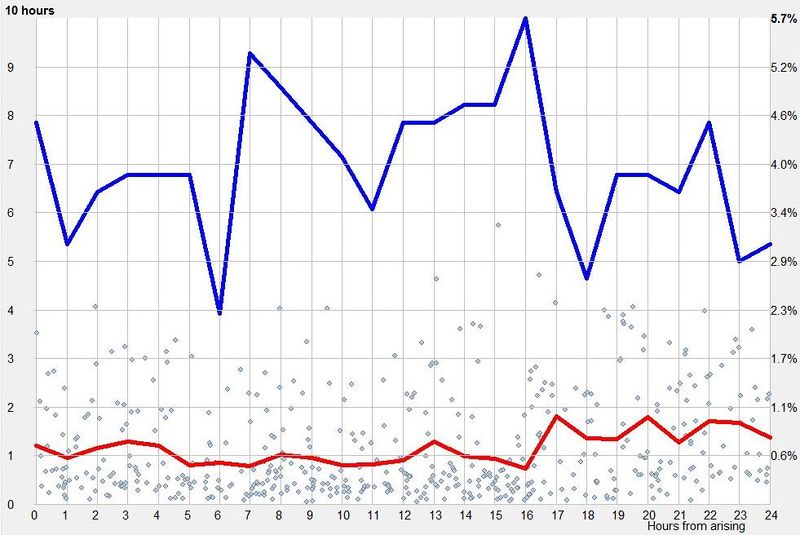
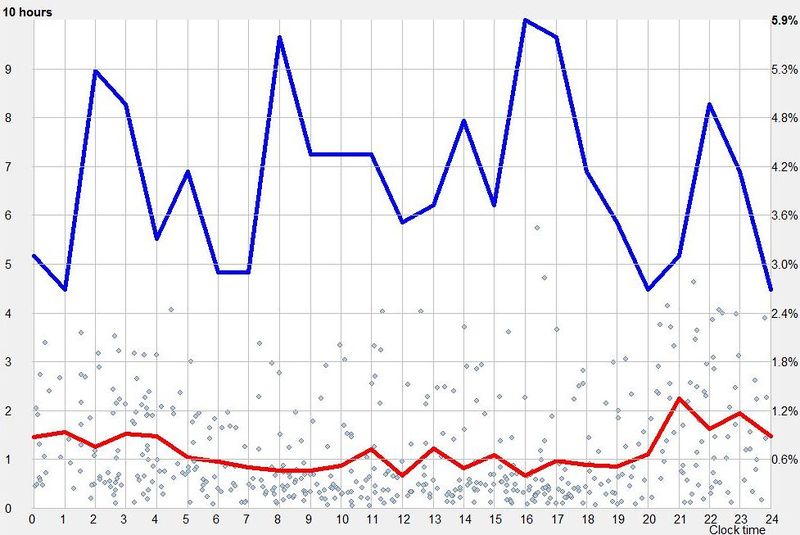
红色昼夜节律线相当平坦,但如第三张图所示,可以通过连续相邻睡眠事件的合并来证明对傍晚和夜间睡眠的一些偏好:
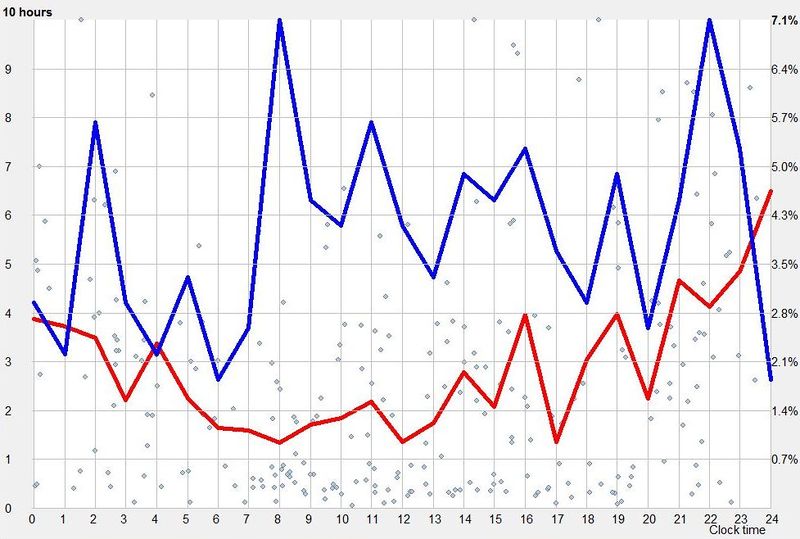
Pirahã 人多相睡眠吗?(Do Piraha people sleep polyphasically?)
虽然科学家不知道任何可以在不损害健康的情况下实际用于缩短睡眠事件长度的自然生物机制,但 Daniel Everett 关于 Pirahã 人的田野报告声称,该部落成员很少每天睡超过 2 小时。我们知道缩短睡眠长度的不健康方式:可以用激素缩短(例如压力);可以使用闹钟(包括基于 ACTH 释放的自然脑钟);可以在错误相位睡眠;可以减少内稳态睡眠驱力(例如用咖啡、毒品、运动等)。所有非自然的缩短睡眠时间方式都会引起睡眠剥夺,其程度是干扰程度的函数。网络上充斥着关于多相睡眠优点的轶事,但没有既定的科学事实可用于断言睡眠长度可以减少。Pirahã 人的例子当然对睡眠科学有吸引力。然而,该部落的难以接近,除了对传教士报告的推测外,几乎没有研究空间。报告可能只是简单的夸大。Pirahã 人也可能是文化主导生理的例子(如”多相睡眠”人群的情况)。我们知晓许多影响昼夜节律周期的突变,可以想象在一个孤立人群中某种特定基因的强流行。然而,这会使 Pirahã 睡眠大大偏离我们灵长类群体中既定的标准。简而言之,如果报告正确,西方人的睡眠习惯将不得不比 Pirahã 部落成员离猩猩更远。
多相睡眠:科学挑战(Polyphasic sleep: scientific challenge)
我不断为关于多相睡眠的科普写作招来批评。然而,这些批评很少触及基本前提,而正是它使预测多相睡眠不能作为合理生活方式选择变得容易。因此,我处于需要忽略批评的阶段,除非它涉及那个基本科学前提:
人类睡眠模式反映潜在的昼夜节律振荡,其周期大致等于 24 小时。人类昼夜节律周期要求每 24 小时一次主要睡眠事件。生物钟可以用最多 3 小时的相位移动来驯化。然而,最大睡眠倾向的昼夜节律周期不能被分割。睡眠倾向顶点的时机不能以相位移动以外的方式定位。周期性不能在不损害健康的情况下被消除。昼夜节律周期是睡眠结构的基础,而睡眠结构对其神经功能至关重要。因此,在睡眠控制系统健康的个体中,没有任何睡眠作息可以绕开主延长夜间睡眠时段。
在实践中,上述前提意味着只有单相和双相睡眠模式是健康且被推荐的。我认为分段睡眠是延长黑暗时段诱发的古老单相睡眠的变体。所有形式的夜间醒来都是常态,应被视为夜间睡眠事件的一部分。在单相或双相睡眠之间的选择取决于昼夜节律波形,它在 24 小时周期内有两个最小值,其中只有一个被证明对健康和福祉至关重要(到目前为止)。
早起者更擅长多相适应吗?(Are early risers better at polyphasic adaptation?)
早起者在多相睡眠中会和猫头鹰一样受苦,时型无关紧要。以昼夜节律周期丧失为特征的不规律睡眠-清醒节律患者,确实在不规律间隔小睡,但他们既不感到精力充沛,也睡得不少于健康个体。早起者、猫头鹰或短睡眠者都无法适应规律的多相作息。多相睡眠可以在警觉性需求高的条件下拯救生命,但它也会缩短被迫实践者的寿命。
为什么对多相睡眠的研究这么少?(Why so little research into polyphasic sleep?)
从轶事证据我可以得出结论,多相睡眠不足以持续造成大量损害。然而,它也延续了大量朗朗上口的迷思,可能影响年轻人对睡眠和整体健康的看法。多相睡眠不是一个整洁的研究对象。科学家喜欢简单,他们构建简单的研究模型以便更容易得出有效结论。我个人喜欢自由睡眠概念作为研究模型,它谈论的是纯净、自然、健康的睡眠。我希望更多研究者更多关注自由睡眠条件,因为所有形式的实验室设计师作息都会在数据中引入一定程度的混乱,常常使数据难以解释或导致错误解释!多相睡眠是为不自然的生存情境提出的,其 Uberman 变体是希望节省睡眠时间或解决睡眠问题的青少年的广泛变异发明。选择多相睡眠作为模型,就像选择一个多行星系统来测试牛顿/开普勒定律,而双行星系统也能做到,并且早 eons 产生结果。不用复杂的傅里叶分析,我们有简单清晰的公式讲述整个故事。
多相睡眠的医疗监督(Medical supervision of polyphasic sleep)
一些多相信徒想知道,在多相睡眠作息下定期去医生处检查以避免潜在健康危害是否有意义。问题在于,家庭医生检测多相作息下麻烦的能力,与他看到新手吸烟者麻烦的能力没有太大不同。损害不是瞬间造成的,也不明显,尽管我相当确信多相睡眠会比吸烟更快造成破坏。认知测试会首先显示变化,可能紧随其后的是免疫功能和葡萄糖代谢。然而,损害很大一部分是多相睡眠的机会成本:不仅是它对健康做了什么,还有一个人本可以因完好心智能力而成就什么。
全科医生无法轻易检测神经层面可能的长期损害,例如睡眠控制系统范围内的。他/她也看不到神经功能变化对患者长期成长和智力成就的影响。你今天不学的东西可能改变你整个生命历程,没有人能估计这个成本。即便是阿尔茨海默病中实质性的神经损害,初期也不易诊断,直到受影响者进入疾病晚期、大脑的相当部分已经消失!人类大脑擅长代偿,发现损害并不容易。
去全科医生那里体检总是一个好主意。然而,作为预防多相睡眠损害的方式,它相当无用。多相睡眠者常报告睡眠剥夺的典型症状:体温调节问题、食欲变化、免疫缺陷等。在短期内很难把这些驱动成严重威胁。毕竟,几小时的”核心睡眠”就能快速补救其中大部分。
有人向我建议:“睡眠剥夺不好,因为它是压力的来源。但它有多糟取决于一个人处理压力的能力。” 确实,睡眠剥夺中对压力的易感性增加,但压力管理是睡眠剥夺的解决方案这一说法是错误的。确实,良好的饮食可能改善吸烟者的健康,但仅靠饮食不能解决吸烟问题。吸烟的唯一终极解决方案是不吸烟。同样,睡眠剥夺的终极解决方案是睡眠。
我自己的多相睡眠试验(My own polyphasic sleep trial)
我常被问到,从未亲身体验如何能声称对多相睡眠有任何权威。首先,我并不自称是多相睡眠专家。作为一个谦逊的生物学家,我只需回忆时间生物学的 ABC 就能得出多相睡眠不可行的结论。你不需要是个瘾君子才能研究药物成瘾,尽管一杯伏特加可能是研究酒精主义的禁欲研究者的推荐一次性治疗。我理解闹钟的痛苦,因为我在大学年代偶尔使用过它。我从早期参与 SuperMemo 业务动荡的岁月中理解时差和睡眠剥夺的痛苦。然而,我的工作需要新鲜的大脑,哪怕一天的模糊头脑都是损失。我无法希望通过挣扎完成多相作息来证明那个难以捉摸、永远遥远的”适应”只是都市传说。如果我要进行自己的睡眠实验,那更可能是分段睡眠的尝试(Wehr,199261)。我可以想象它对学习和创造力有奇效。然而,这个世界上很少有人能负担 10 小时的清醒日。似乎只有带薪志愿者愿意品尝过度睡眠的福祉。在肤浅的读者从 Wehr 的工作得出多相睡眠可能的结论之前,让我强调,他的分段睡眠实验谈论的是非常长的睡眠块,而非 Uberman 式的迷你小睡。
绘制多相睡眠(Charting polyphasic sleep)
数据来源(Source of data)
在发表《Polyphasic sleep: Facts and Myths》(Wozniak,200562)后,几十位年轻男子写信给我,请求协助驯化多相睡眠作息。出于伦理,我只能从尝试劝阻年轻热情者继续实验开始。毋庸多说,这些都不是容易被说服改变方向的人。正如我在《Facts and Myths》中所写,这些是”准备寻求最大生产力新方式的叛逆者”。在这种情况下,没有任何科学论证足够有说服力,毕竟所有推理都很容易被”科学还没有所有答案”压制。没有一个年轻叛逆者成功驯化多相睡眠,然而一些人足够坚持,提供了有价值的 SleepChart 数据,帮助阐明多相睡眠作息长期使用的不可行性。
Stampi 帆船研究(Stampi yachting research)
在Stampi 获得的数据中,我们看到一位单人水手在实际帆船比赛中半多相睡眠的时机。在这个案例中,昼夜节律图揭示了清醒日前半部分的睡眠禁区,以及在清醒日 15-24 小时启动睡眠的清晰昼夜节律偏好(蓝线):
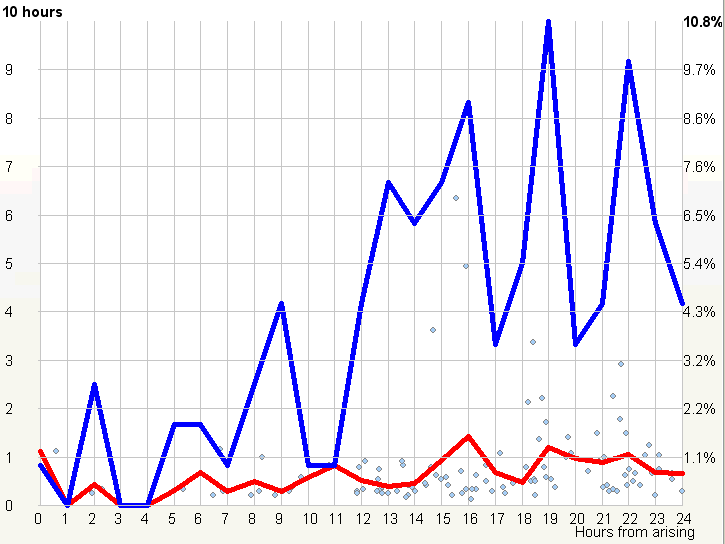
由于睡眠被人为打断,红色昼夜节律曲线在这里没有意义。此外,人为控制睡眠是睡眠维持曲线与睡眠启动曲线之间角色反转的原因。在这种情况下,是睡眠启动曲线最佳地表达了昼夜节律睡眠倾向。
为这个看似嘈杂的睡眠生成的周期图显示了典型的双相模式,峰值在 23.9 小时和 12.1 小时。23.9 小时的日和相关的相位前移最可能由向东航行时跨时区的影响造成:
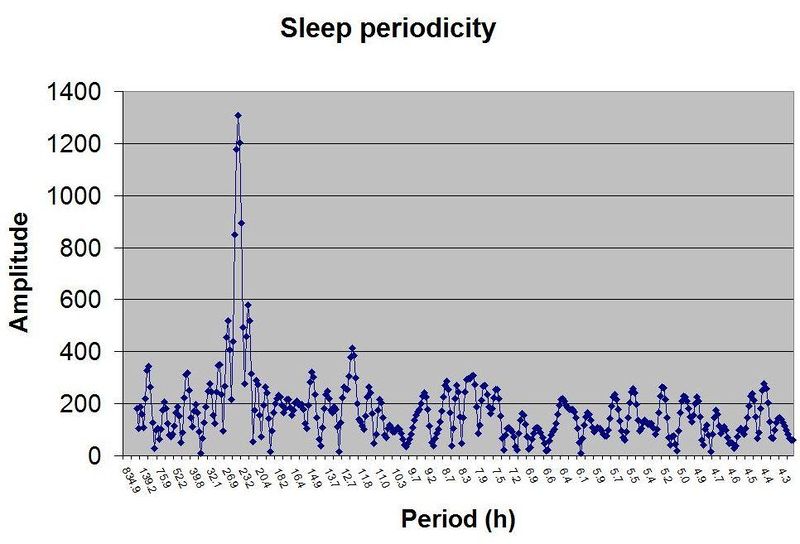
多相睡眠尝试(Polyphasic sleep attempt)
在试图驯化多相睡眠作息的尝试中,一位男性信徒以 4 次 30 分钟小睡和 20:00 的 3 小时”核心睡眠”的作息开始实验,打算在”适当时候”把它减少到 30 分钟。驯化(“适应”)似乎难以捉摸,因为信徒不断在一些小睡时无法入睡,同时在一些分配的清醒时段继续与警觉性斗争。昼夜节律图显示超日节律睡眠启动,对在主观日开始后第 14-22 小时主观夜晚时段睡眠有昼夜节律偏好:
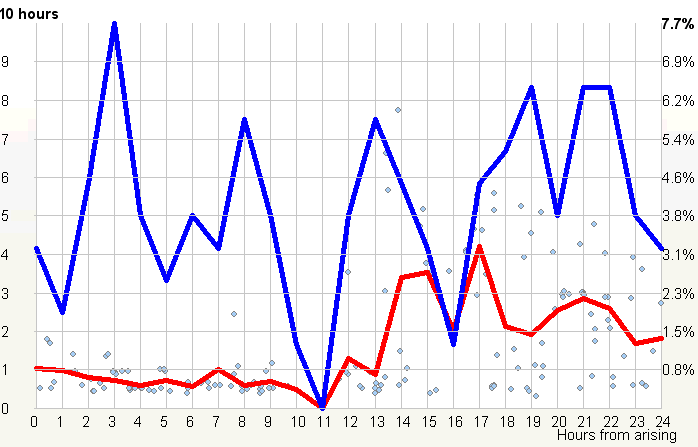
核心睡眠无法如计划那样缩短,否则会进展到越来越严重的睡眠剥夺。相反,核心睡眠长度略有增加,并移到更晚时段。逐渐地,白天小睡开始消失,直到信徒移到典型的双相睡眠:傍晚 5-6 小时,加上早上 30-60 分钟小睡(如果核心睡眠导致严重睡眠剥夺,白天偶尔有额外小睡)。一年后,该信徒接近单相,只剩下一条规则:“尽量在午夜前入睡”。这项努力记录在这个博客中。
Uberman 睡眠日志(Uberman sleep logs)
图片显示了我从潜在多相睡眠者那里收集到的三份最自律的 Uberman 睡眠适应尝试的 SleepChart 日志。该图说明了 Greg(A)、Bryan(B)和 Claudiu(C)的努力:

Greg 的尝试在 9 天后被实验后期清晨时段”核心”睡眠的聚集所挫败(日志 A)。这种聚集无疑是由不断累积的睡眠剥夺加上主观夜晚时段昼夜节律睡眠倾向的峰值造成的。
Bryan 的尝试总长 22 天,其中 13 天没有”核心睡眠”,是最长的(日志 B)(完整日志包含在下一节)。这次尝试第一天就显示出”紧张”迹象,头四天有四次额外小睡。一些睡过头在第 7 天开始在主观夜晚时段显示合并。最后,第 14 天中午,受试者陷入了一段漫长的 11 小时恢复性睡眠。该尝试又继续了约 8 天,伴随多次额外小睡、睡过头、昏沉与亢奋交替的时段。最终,Bryan 的详细笔记得出了一个有趣的结论:睡眠倾向的昼夜节律周期很可能在整个实验期间在后台自由运行,显示近乎完美的 25 小时周期。详细分析和解释见:多相睡眠中自由运行的昼夜节律周期。
Claudiu 的尝试在经历第一次额外小睡失守前持续了近 5 个整天,是”纯 Uberman”中最长的(日志 C)。它同样值得注意的是,第一天之后从未错过任何一次小睡。然而,重要的是要注意,许多小睡者发现很难确定,在禁区到来的小睡中他们是否真的睡着了。他们标记为小睡的可能实际上只是几个短暂的微睡瞬间。
大多数声称在多相睡眠上成功的博主似乎都降低了他们对满意警觉性和创造力的标准。当”我成功的实验”和”昏沉”这样的陈述同时出现时,你可以确信”实验”并未有效最大化他们的警觉性和生产力。睡眠惰性对健康睡眠者应是陌生的。
博客圈中相当多的”成功”Uberman 睡眠声明可以有多种解释。然而,这些”成功”没有一个可能用使多相睡眠如此难熬的自然昼夜节律性的消失来解释。如果信徒确实变得无节律,这将是严重的健康和长寿风险。在最好情况下,这可能意味着睡眠控制中心的失调和解耦(见:Sleep-wake flip-flop)。在最坏情况下,这可能涉及对负责睡眠控制的脑中心的胶质损伤。毋庸多说,这样的失调或损伤变化难以逆转,并会导致获得令人神清气爽睡眠的严重问题,那将是 Uberman 希望者目标的对立面。
双过程睡眠模型 vs. 多相睡眠(Two-process sleep model vs. polyphasic sleep)
从 Greg(A)、Bryan(B)和 Claudiu(C)收集的多相睡眠数据(见前文)用 SuperMemo 处理,试图了解在不可预测的昼夜节律相位下小睡时机如何影响睡眠倾向:
SuperMemo 实现了 Borbély 模型双过程模型的变体,使得基于睡眠和清醒历史预测警觉性和/或睡眠倾向成为可能。SuperMemo 15(或更高版本)的用户可以在时间线任意选定点上 Shift+click 睡眠日志来检查自己的睡眠倾向预测数据。
在所示的图中,粗红线代表估计的警觉性,即睡眠倾向的倒数。昼夜节律睡眠倾向以青色标记。很容易看出,警觉性曲线的形状取决于小睡发生的昼夜节律相位。这些图被并排放置,以便在小睡发生在产生不同警觉性估计的不同昼夜节律相位时对齐小睡时机。这些图表明,同样的小睡时机可能产生完全不同的警觉性曲线。这解释了为什么多相睡眠的”能量化力量”是一个容易传播的迷思。
在多相睡眠中,根据昼夜节律相位,小睡可能产生高度警觉性或严重的睡眠惰性。
尽管 SleepChart 中使用的睡眠模型适用于自由睡眠,Uberman 小睡的对称性使得正确预测昼夜节律峰谷变得不必要。无论顶点峰值出现在哪里,它们都会在很大程度上随机地与小睡网格相交。此外,如果在多相睡眠实验前收集了规律睡眠数据,该时期正确的客观夜间顶点估计将延续到多相实验的头几天。毕竟,只有一致的相位移动才能重新定位昼夜节律正弦波的相位。使多相睡眠难以维持的同样机制,也可以用为自由睡眠条件设计的模型来解释。
多相过山车(Polyphasic rollercoaster)
把双组分模型应用于上述”Uberman 睡眠”数据,主要浮现的是警觉性和昏沉交替的典型”过山车效应”。与醒来神清气爽、入睡疲倦的典型睡眠者不同,多相睡眠者会经历极度亢奋(如图 B 中凌晨 4 点)和令人沮丧的低谷(如图 C 中下午 3 点)。所示的警觉性估计与睡眠者自己的主观”专注和动机”评估相关良好。在图 C 中,晚上 11 点的小睡产生了警觉性的主要激增,而仅仅 4 小时后凌晨 3 点的小睡几乎什么也没带来。这在多相睡眠者中产生了热情与自我怀疑的典型过山车。在昼夜节律睡眠倾向最小值处发生的短暂小睡后,多相睡眠者可能达到普通睡眠者未知的高度。那些接近亢奋的热情激增相当独特,因为普通睡眠者几乎从不在昼夜节律睡眠倾向最小值处小睡。这些时刻可能让多相实验者更新博客:“从未感觉这么好——创造力达到最大”。同时,一些小睡只会让事情更糟。例如,图 C 中 2009 年 6 月 23 日下午 3 点的小睡似乎没有产生任何警觉性提升,随后是一小时长的”修正”,同样没有提升警觉性。此外,达到峰值的昼夜节律波总是由于昼夜节律睡眠惰性产生令人不快的昏沉感。这是轮班工作者相当熟悉的惰性类型,也是时差大脑模糊的原因。这说明了 Stampi 博士在实验中注意到的:在多相睡眠作息下保持清醒不难,多相作息最难的部分是从昼夜节律顶点处的小睡中醒来的过程(Stampi,199253)。
多相睡眠中自由运行的昼夜节律周期(Free running circadian cycle in polyphasic sleep)
Bryan 的多相尝试(前文提到)带来了最有趣的观察:昼夜节律周期很可能在后台运行,就像在无授时因子条件下一样。与多相睡眠事件对睡眠控制系统的混乱影响一致(见:Phase response curve in polyphasic sleep),这些事件在很大程度上互相抵消,允许一个自由运行的昼夜节律周期在后台运行。此外,如果选择正确的睡眠相位,回到单相生活方式可以几乎无痛。
Bryan 在 Excel 中保存日志,带有详细注释。有趣的是,他用颜色表示警觉或甚至亢奋的时段,以及昏沉或困倦的时段(黄色):
睡眠
持续的困倦、疲劳、缺乏专注力与动力
正常/中性状态,不过度困倦也不过度清醒
高度的头脑清晰、专注、动力、热情
列对应实验的连续日子,包括之前和之后的单相睡眠区块。行对应半小时时段。睡眠区块用其以分钟为单位的持续时间标记。在图中,Bryan 的 Excel 笔记被稍作修改,以可视化主观夜晚的范围。最重要的是,他的睡过头区块被标记为黄色而非蓝色,以区别于人为强加的多相睡眠(后者不告诉我们他实际的睡眠倾向)。这样,一条视觉上明显的对角黄色带出现了,那里推测的最大昼夜节律睡眠倾向被标记,没有区分睡过头事件和昏沉时段。我用蓝线和红线的就寝和醒来线(主观夜晚的假定边界)划定了那条黄色带,方式与 SleepChart 中相同。这些线的角度以及相关的昼夜节律周期相位移动指向近乎完美的 23 天回转,大致对应 25 小时的生物钟周期。换言之,我们可以猜测,尽管有多个混乱且不可预测的输入到相位移动系统,昼夜节律周期仍在后台自由运行。同样值得注意的是,Bryan 通过命中其自由运行昼夜节律周期中主观夜晚的确切时段,多么容易地回到了单相生活方式。
Bryan 的 SleepChart 日志显示,SleepChart 1.0 中使用的划定主观夜晚边界的统计方法未能追踪他假设的自由运行昼夜节律周期:
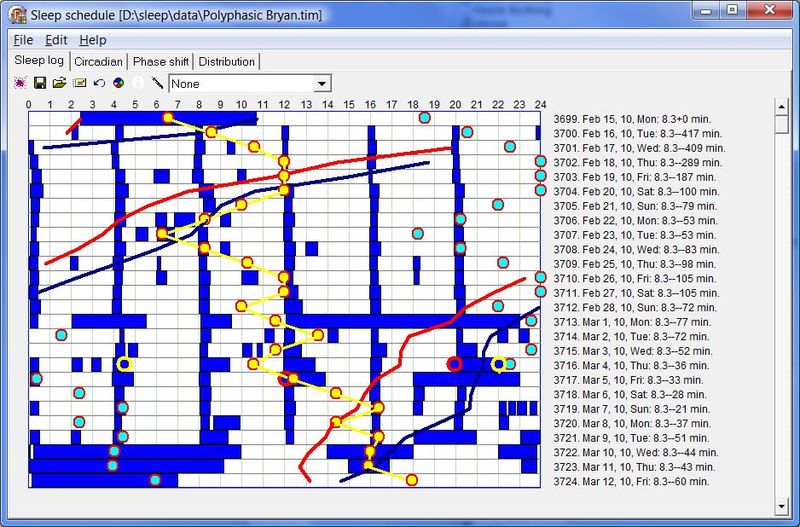
图中的红线和蓝线显示相位前移,而节律几乎肯定显示相位延迟。失败的原因是 SleepChart 1.0 的统计方法使用睡眠区块作为睡眠倾向的标记。自然,在多相睡眠中,这些标记被伪造,使算法陷入混乱。另一方面,基于相位响应曲线(PRC)的 SleepChart 2.0 中使用的较新方法,能够大致追踪 Bryan Excel 文件中注意到的昼夜节律低谷。在这里,黄色昼夜节律峰值线在 2 月 20 日至 23 日期间有些混乱,这就是为什么近似未能在时间上恢复以匹配回到单相睡眠的原因。
尽管 Bryan 充其量只能连续维持纯 Uberman 作息 3 天,但他一个月的努力仍是自律的显著展示。如果你读 Bryan 自己关于认知功能的笔记,你可能会同意,多次困倦、疲劳或昏沉的时段,使多相睡眠不成为靠脑力为生者的生活方式选择。然而,Bryan 自己总结实验的话相当令人惊讶。他似乎不被”困倦时段”或”醒来困难”所困扰,而这些绝不应是管理良好、卫生的睡眠模式的一部分:
“我的实验向我明确无误地证明,在多相睡眠作息下维持正常(主观)功能(心理和身体)是可能的,如果你愿意适应严格的小睡作息,并忍受每天持续 2-4 小时的困倦(昼夜节律低谷)时段。与 Stampi 的发现并行,我经历的唯一重大困难是随着实验推进从小睡中醒来。醒来变得越来越困难;有时我会醒来并在没有任何记忆的情况下重置闹钟;或者我女朋友不得不摇晃我一整分钟直到我醒来。然而,一旦醒来,我通常感觉很好,就像睡了一整夜——但没有在床上躺 8-10 小时的宿醉(昏沉)。把注意力集中在醒来的困难上、声称这表明什么或不表明什么很诱人,但清醒时段正常甚至亢奋的功能不应被忽视。鉴于此,我很高兴看到你在题为 Polyphasic rollercoaster 的段落中讨论了这些事情。”
很可能在多相睡眠尝试中,昼夜节律周期在后台运行,就像在无授时因子条件下,混乱的相位移动输入互相抵消。
Claudio Stampi
可能没有人比 Claudio Stampi 博士更了解多相睡眠。他毕生致力于理解超日节律和小睡艺术。他对这个想法的热情源于三十年前,当时他还是医学生,同时也是一位热忱的单人水手。他研究了数十位参加竞赛航行者的睡眠,为 NASA 研究睡眠模式,并在实验室条件下研究多相睡眠。他用腕戴式活动监测仪和脑电图电极给受试者佩戴。他是小睡的崇拜者,因为没有什么比小睡更能对抗睡眠剥夺和疲劳。在他的工作中,他寻找提高警觉性和在危及生命情境(尤其是长距离帆船赛)中生存的方法。然而,他不向能负担一整夜健康睡眠的正常运作的创造型个体推荐多相作息。他所谓的”推荐”只是与多相睡眠模因一起流传的迷思之一。使用多导睡眠图工具,Stampi 寻找人类警觉性的低谷和峰值。他的研究试图利用对这些超日节律的理解,主要通过优化小睡时机来最大化小睡的有效性。
Stampi 的方法主要针对最小化睡眠剥夺。他自己是个双相睡眠者,凭借其时间生物学专业知识可以自豪地声称*“我从不疲倦”63。谈到 Ellen MacArthur64 时,他用一句话概括了他的研究:“Ellen 所做的,是在她睡眠的需求和一直保持清醒的需求之间找到最佳折衷。“*
多相 vs. 创造性生活(Polyphasic vs. creative lifestyle)
与单人水手不同,创造型个体不需要折衷。追求的是警觉性、注意力和创造力的不妥协最大值。Stampi 已经证明,与单相睡眠相比,多相睡眠在睡眠剥夺条件下可以改善认知表现:“每 4 小时睡 30 分钟、每天总共只睡 3 小时的个体,表现更好、更警觉,相比他们连续睡 3 小时。换言之,在大幅减少睡眠的条件下,更频繁地给睡眠’电池’充电更高效。” 许多人把这用作多相睡眠优越性的论据,同时悄悄绕过 Stampi 也注意到多相作息的表现仍远低于自由睡眠条件下的事实。
多相圣经(Polyphasic Bible)
多相睡眠的许多支持者会引用他们的圣经:Claudio Stampi 20 多年前写的《Why We Nap: Evolution, Chronobiology, and Functions of Polyphasic and Ultrashort Sleep》。Why We Nap 是一本优秀的书,充满关于睡眠、小睡、演化等的珍贵信息片段。我可以全心全意推荐这本书,作为该领域最权威专家有趣文本的绝佳汇编。书中还包括关于列奥纳多·达芬奇假定睡眠习惯的轶事笔记。正是这个轶事的纳入可能产生了意想不到的副作用,导致了 Uberman 大爆炸。凭借额外二十多年睡眠研究的优势(例如昼夜节律周期的基因方面),我不同意 Stampi 的一些原始假设,Stampi 自己在很大程度上也不同意。这不改变他的研究绝对可被视为研究时间生物学(chronobiology)极限的先驱工作这一事实。对于那些仍相信 Stampi 倡导多相睡眠作为生活方式的人,他书中的一个古老引用应该能澄清事实:“作者想提醒不要对这些结论进行误导性解读。这里提出的不是多相睡眠优于单相睡眠,也不是每个人都应该转向多小睡行为’万灵药’。显然,准单相睡眠——单相睡眠加偶尔小睡——对大多数成年人和少数其他物种来说最自然。如果在演化中的某个地方,这样的物种发展出维持相对长时间清醒的能力,这种能力最可能是对某种重要且有利的适应压力的响应。”。
Stampi 博士写道:“许多实验提供了直接证据,证明成年人对多相睡眠-清醒行为的不同类型——和不同水平——有令人惊讶的适应能力。” 这一陈述足够笼统因而是正确的。例如,睡眠阶段压缩是一种适应形式。这并不意味着个体能在一天中多次自然小睡。在睡眠研究中形成共识:在健康成年个体中,多次爱迪生式小睡需要一定程度的睡眠剥夺。没有剥夺,启动睡眠变得相当困难。“多次小睡”应被理解为”合并后超过一次”,其中”合并”是把间隔很近的多次小算作一次的过程。
多相睡眠中的认知测试(Cognitive tests in polyphasic sleep)
重要的是要注意,Stampi 博士在其多相睡眠实验中只能识别认知功能的轻微下降。这似乎与其他研究或简单的记忆表现昼夜节律测量(包括用 SuperMemo 可能进行的测量)相矛盾。在测量中纳入昼夜节律组分,会在正常不剥夺的清醒日中产生显著的认知差异。简单的记忆测试,如果取平均,可能因过山车效应产生看似合理的认知表现评估。Stampi 选择测量认知表现的测试,回避了最佳时机 REM-NREM 交互在睡眠中主要长期神经生理功能这一本质问题。在 Stampi 对 Francesco Jost 的实验中,REM 和 NREM 很少同时发生。如果考虑假设的记忆存储优化功能,无法用短期测试验证记忆状态。这是因为,理论上,把大脑网络功能视为黑箱应保持不变。对睡眠结构的忽视只会表现为大脑建立新技能和推理能力的累积性长期无能为力。其次,优化存储的创造潜力也难以测量,且肯定显示需要长期研究的累积效应。最后,缺乏昼夜节律效应只能证明所选测试的敏感性和/或时机不足。即便内稳态警觉性组分确保我们看似能专注于简单的心理任务并执行得相当好(如记忆任务、驾驶、简单计算等),昼夜节律低谷也会影响维持脑力努力的能力或削弱其创造性方面。对 Stampi 的目标(如最大化单人帆船赛中的警觉性)足够的测试,不能用于声称超短睡眠对认知表现的长期影响。
睡眠科学文献中,在睡眠的内稳态和昼夜节律组分及其对认知的影响之间存在一定程度的混淆。研究者研究环境因素对睡眠的影响时,常常未能区分两者。我们都知道咖啡能帮助度过困倦时刻。然而,重要的是问,它的效果是内稳态的还是昼夜节律的?咖啡能驱散睡眠惰性吗?能帮助克服昼夜节律低谷吗?如果咖啡对昼夜节律困倦的影响可以忽略,仅仅说咖啡帮助克服困倦是不够的。任何熟悉时差的人都能证明,大脑模糊的状态并不明显剥夺人的基本心理技能,然而它可以通过影响自我控制、创造力、动机等完全毁掉一个人的生产力。这就是为什么环球奔波的政治家是开创性和平或贸易协议的糟糕材料,即便他们相信能在 3 小时睡眠或时差状态下运作良好。Stampi 博士的发现(高度适用于紧急情况)不应用于削弱时机恰当的自然睡眠对大脑功能的重要性,以及人为设计师睡眠作息非常有害这一事实。
睡眠剥夺就像醉酒(Sleep deprivation is like alcohol intoxication)
睡眠研究者喜欢把睡眠剥夺比作醉酒:两者都破坏自我评估能力。就像酗酒者总声称”我没醉,我只是微醺”,睡眠剥夺者常说”我很好,我敏锐警觉”,而他/她执行心理任务的能力可能严重受损。人越困,对自己执行认知任务的能力越过度自信。睡眠剥夺驾驶可能与醉酒驾驶一样危险。这种自我评估能力的丧失,可能部分解释了为什么那么多多相睡眠博主声称他们已适应了严苛的作息。他们倾向于在清醒和/或亢奋的时刻写下成功(见多相过山车)。同时,他们把那些脑死亡时刻当作”暂时挫折”、过渡适应状态等而忽略。在那些模糊时刻,博主可能不愿更新博客,放大了其报告认知中的偏差。自然适应多相作息是不可能的,但那些夸耀声称者不必被贴上骗子标签。自我评估障碍和期望标准的降低,都应作为减轻情节。如前所述,甚至可能把昼夜节律功能扁平化或解耦到足以减轻主观夜晚时段醒来的痛苦。由于这种缓解来自功能失调,甚至可能来自神经损伤,它不应给那些希望适应的人带来安慰。鉴于所有遗传级联都依赖昼夜节律周期,这样的结果只会导致健康灾难。
睡眠债与小睡(Sleep debt and napping)
PureDoxyk 是”Uberman 睡眠作息”的”发明者”的昵称。尽管她声称多相睡眠了较长一段时间,但更仔细地审视她的报告表明,她以一种”混乱的多小睡代偿性睡眠系统”睡觉,逐渐向相当自然的双相睡眠漂移,她后来称之为”Everyman 睡眠作息”。如果不是这种漂移和采取”核心睡眠”的倾向,我甚至可能怀疑 Uberman 睡眠周期的发明者患有导致昼夜节律无节律性的罕见突变。患有这种障碍的人无法在夜间长时间区块睡眠,而在白天多次小睡。这些小睡加起来达到相当正常的总睡眠时长,并产生相当不神清气爽的头脑,使该障碍相当难以生活。如果是一个饱受糟糕睡眠之苦的病人提出了一个睡眠”系统”,结果导致青少年为寻求更好睡眠而进行生活方式实验的流行病,却只找到更多睡眠时间的痛苦,那将是一个讽刺而悲伤的转折。
PureDoxyk 定律(PureDoxyk Law)
PureDoxyk 的文字令我印象深刻的是,她立即显得可信,似乎对睡眠剥夺与小睡之间的联系有非常好的感觉。让我们看看她的声明,我称之为 PureDoxyk 定律。注意”六小时睡眠”片段,它表明 PureDoxyk 没有像上面推测的那样患有严重的昼夜节律无节律性障碍:
六次小睡无睡眠;四次小睡一个半小时睡眠;三次小睡三小时睡眠;两次小睡四个半小时睡眠;一次小睡六小时睡眠。
* 我从定律中移除了两个微小的数学扭结,原定律表述为:六次小睡无睡眠;4-5 次小睡一个半小时睡眠;3 次小睡三小时睡眠;1-2 次小睡四个半小时睡眠;一次小睡六小时睡眠(来源)。
显然,这个定律需要参数化以适合一般健康人群。尤其是,大多数单相睡眠者会发现每天难以小睡超过一次,除非所有睡眠事件都被闹钟终止,从而使睡眠剥夺的循环延续。
我们可以立即看到夜间睡眠持续时间与所采取的小睡次数之间近乎完美的线性关系:
NapNumber = 5.6 - 0.8 × CoreSleep
最小化总睡眠时间(Minimizing total sleep time)
如果 PureDoxyk 定律成立,小睡时长将决定多相睡眠中整体时间收益的盈亏平衡点。超过盈亏平衡点的小睡时长,增加额外小睡会增加睡眠的总成本。显然,盈亏平衡点将与总睡眠量恒定(即与小睡次数无关)的情况重合。如果我们取总睡眠为:
TotalSleep = CoreSleep + NapNumber × NapDuration
从 PureDoxyk 定律代入 NapNumber,对小睡时长微分,并把结果与零比较,我们将得出 NapDuration = 75(分钟)的盈亏平衡点,对应恒定的 7 小时总睡眠时间。换言之,添加少于 75 分钟的小睡会在多相睡眠中产生整体时间收益。
一张理论图,显示沿 PureDoxyk 定律最小化总睡眠时间。近端水平 X 轴显示小睡次数,后退水平 Y 轴显示小睡时长,垂直 Z 轴显示以小时为单位的总睡眠时间。盈亏平衡小睡时长线标记为”75 min”。该图显示,添加少于 75 分钟的小睡可以实现总时间收益,而添加超过 75 分钟的小睡将增加总睡眠时间。
分析符合上述定律的不规律睡眠日志将很有趣,因为它们可以回答一些关于自由睡眠调节力量与多相睡眠中使用闹钟诱发的睡眠压缩适应力量之间睡眠效率拔河胜者的问题。
短小睡作息下的净时间收益显然不会转化为大脑收益,这不应被理解为寻求最小总睡眠时间的建议。我提出上述问题只是作为一个有趣的数学关系,它为总睡眠债提供了一个整洁的公式,可能在建模睡眠被过早终止(例如用闹钟)条件下的睡眠时有用。SleepChart 和 SuperMemo 都不考虑睡眠债,因为两者都是为理想的自由睡眠条件设计的。显然,任何形式的睡眠债都是不受欢迎的,因为它意味着未完成的睡眠神经功能。
与其追求最小化睡眠时间,我们应追求最大化睡眠的大脑效应。
最佳小睡时长(Optimum nap duration)
当夜间核心睡眠时长与总小睡时长之间的实际相关性被研究时,出现了一个相当非线性的关系:
在本例中,负指数函数对数据的拟合远好于线性函数。然而,在与 4-8 小时夜间睡眠相对应的最受研究范围内,可以观察到近乎线性的关系:每丢失一小时夜间睡眠需要 20 分钟的替代小睡时间。这表明小睡具有强大的代偿能力。
然后我们可以重新表述 PureDoxyk 定律,使其适用于更广泛的人群。最重要的是,一次中午小睡应足以代偿所有丢失的夜间睡眠(见:Best nap timing 和 One nap per day is enough)。因此,用一次时长取决于丢失睡眠量的小睡取代多次小睡更有意义:
NapDuration = (SleepRequired - SleepObtained) / 3
这个公式只对时机恰当的小睡成立。过早的小睡不能提供完全代偿。过晚的小睡会更长,并缩短次夜的睡眠。
在睡眠剥夺条件下,夜间睡眠债需要额外小睡时间,大致呈 3:1 比例。每丢失一小时夜间睡眠,需要额外 20 分钟小睡。
同样,这个公式应只具有理论价值。你绝不应试图终止代偿性小睡。如果时机恰当,它应被允许走完自然进程,然后它将为丢失的夜间睡眠提供最佳代偿。
尽管小睡为丢失的夜间睡眠提供了绝佳代偿,它们无法提供完全的功能替代。要达到最大认知能力,你需要不间断地运行夜间睡眠直到完成!
不规律睡眠者的人格特征(Personality characteristics of irregular sleepers)
PureDoxyk 在没有任何专门记录工具的情况下获得了足够的睡眠多相经验来阐述上述定律,这表明她需要相当广泛的小睡排列组合才能看清大局,而在此案例中,这个大局似乎非常合理。PureDoxyk 定律可以被解释为健康的单相或双相睡眠如何随着睡眠债程度的增加被拉伸到多相睡眠相空间的演示。PureDoxyk 自己把她包含”核心小睡”的新睡眠作息称为 3 小时 Everyman 作息。如果不过度依赖闹钟,这种作息听起来相当可持续。毕竟,三分之一的美国人尽管犯下每天平均按 3 次贪睡按钮的日常神经罪行,仍能相当正常地运作。毋庸多言,这种 Everyman 作息与我最初觉得特别麻烦的 Uberman 原始表述相去甚远。
过去,我收到了一些睡眠模式相当不规律(包括多次小睡)的睡眠日志。这些日志伴随着一些轶事证据,似乎表明那些不规律模式与某些人格特征强烈相关。我在这里可以广泛推测,说那些是相当神经质但又相当有创造力的类型(排除可归因于处方药使用的案例)。如果情况如此,那些睡眠模式可能对长寿不太好,但即便自由睡眠也无法把它们理顺。这表明可能有基因因素涉入,而”突变假设”远更可能解释持续的不规律模式,而非规律的神清气爽 Uberman 模式。我甚至在这里会避免使用”突变”一词,因为这些”人格基因”在人口中必定相当普遍。PureDoxyk 的案例如何解释我毫无头绪,但它似乎在其独特性上并不太极端,因此,从概率上讲,它可以被认为是可信的。
多相天才(Polyphasic geniuses)
网络传言,有许多天才多相睡眠。言下之意是,如果多相睡眠对历史上最伟大的头脑有效,它也应该对一个渴望征服世界的年轻有抱负的学生有效。然而仔细审视,那些多相故事非常难以证实。不知何故,这一群体不包括当代诺贝尔奖得主、总统或伟大运动员。换言之,你不能只是给名人发邮件问。所有伟大的多相睡眠者都已去世。仍有几位在博客中夸耀自己是多相睡眠者。他们的睡眠常常只是双相睡眠定义的延伸,或是各种睡眠模式与大量睡眠剥夺的组合。其中一些案例,我只能用既得利益或膨胀的自我来解释。由于他们的”成功”晚于 Uberman 睡眠作息的”发明”,这可能只是一个希望被加入伟大 Übermenschen 名单的、永远不可信的愿望。即便发作性睡病也无法解释一些多相信徒的小睡习惯。无论如何,成功的多相睡眠案例无论如何无法验证。自然,缺乏证据并非不存在的证据,本节并非旨在证明多相睡眠不可能。是上面的生物学论证解决了问题。这里,我只是试图说明互联网和人性的造迷力量。
随着 22 小时清醒日益处的雪球般迷思,多相天才的名单似乎越来越长。名单包括达芬奇、爱迪生、特斯拉、丘吉尔、本杰明·富兰克林、托马斯·杰斐逊,甚至李小龙。如果牛顿和亚里士多德很快加入我也不会惊讶,也许耶稣随后也会跟上。
我试图找出历史上最伟大天才的睡眠习惯是否有任何记录。我能找到的,无非是对正常单相或双相的标准遵守,在创造性高峰时期有无数次通宵的例外。
巴克敏斯特·富勒(Buckminster Fuller)
在 Buckminster Fuller 这里,我最接近找到一种准多相作息。我设法联系上的 Buck 传记作者证实,他的睡眠习惯相当不寻常,他试验了各种睡眠模式。尤其是,在广泛旅行和讲学期间,他会进入他所谓的”狗睡(dog sleep)“。然而,那种睡眠与多相睡眠毫无关系。它是自由睡眠与时差、会议和截止日期混合的即兴组合。换言之,Bucky 会在疲倦且有机会时打盹。然而,如果能在这里或那里挤出扎实的 6 小时,他不会错过机会。这种”狗睡”不符合任何固定的闹钟作息,只是昼夜节律与 Bucky 忙碌生活方式之间的折衷。
列奥纳多·达芬奇(Leonardo da Vinci)
尽管连 Stampi 也轶事性地提到 列奥纳多·达芬奇,但列奥纳多的多相睡眠很可能是都市传说。我找不到任何关于他睡眠习惯的可信来源,然而每当小睡艺术的问题出现时,几乎总会提到达芬奇。似乎很奇怪,在以阳光优于烛光为计时方式的悠闲文艺复兴生活中,有人会想出疯狂的多相作息主意。据说,他暗示了单相心态,谈到死亡:“正如充实的一天带来快乐的睡眠,善用的一生带来快乐的死亡。” 更说明问题的是 Bandello 关于达芬奇创作《最后的晚餐》的报道。列奥纳多会从黎明到黄昏连续工作,忘记吃喝。震惊的 Bandello 肯定会报道一位多相睡眠者通宵工作更为惊人的事。在我看来,用 500 年前一个研究不充分的历史案例来支持多相睡眠,更像是过时的**诉诸权威(argumentum ad verecundiam)**。
我怀疑整个列奥纳多迷思可能源自一位灵媒 50 年前讲的故事!Giancarlo Sbragia 在他关于超短睡眠的文本(1992)中报告:我记不清从哪里或从谁那里收集到关于列奥纳多睡眠习惯的信息。[……] 我有一个朋友是灵媒,能超感知觉。[……] 很可能是从她那里我了解到列奥纳多奇特的睡眠-清醒模式,尽管 30 年后的今天,我并不完全确定。(Sbragia,199265)。
顺便说一句,达芬奇也是出现在许多其他可疑名单上的名字:患有注意力缺陷障碍的伟人名单,或伟大素食者名单。他也是都灵裹尸布的可疑伪造者。同样的模因机制必定反复地把达芬奇、耶稣、爱因斯坦、爱迪生、杰斐逊、富兰克林和希特勒放在许多迷思中。他们不断出现在关于受 X 影响、实践 Y 或相信 Z 的名人的捏造名单上。
拿破仑·波拿巴(Napoleon Bonaparte)
拿破仑在小睡或多相睡眠的背景下被提及的频率不亚于达芬奇。而他的案例通过历史记录相当容易证伪。与列奥纳多的艺术天才相比,一位杰出的军事指挥官在持久战中或在他充满无数事务的紧张生活中退休小睡,似乎更加荒谬。据说他确实睡得少且频繁。他在巨大压力时期患失眠,也常被信使打断,这可能增加他白天小睡的倾向。然而,他只在有坏消息时被叫醒,硬性规定好消息可以等。他的回忆录表明他不介意早死,因此他会无视医生关于少睡和大量喝浓咖啡的建议。由于拿破仑的生活充满压力,他的短睡眠可能是其生活方式的结果。低睡眠饮食对拿破仑的军事技能转化不佳。一些同时代人把他滑铁卢的失误归因于睡眠剥夺。然而,在较慢的日子里他会睡扎实的 7 小时,7 点醒来常常赖床到 8 点,然后在下午再加一次小睡。记录还表明,在圣赫勒拿岛他是一个正常的睡眠者,当压力被无聊取代时,他常常睡到很晚。
托马斯·杰斐逊(Thomas Jefferson)
杰斐逊似乎也容易作为多相睡眠者被证伪。在给 Vine Utley 医生的信(1819)中,托马斯·杰斐逊写到他的睡眠习惯。我们可以得出结论,他的睡眠不太规律,他会在不同时间入睡(常常到深夜),睡前总是至少花 30 分钟进行创造性阅读,如果阅读特别有趣会更晚入睡,并规律地在日出时醒来。换言之,不出所料,杰斐逊的生活中没有多相睡眠的痕迹。
本杰明·富兰克林(Benjamin Franklin)
至于 本杰明·富兰克林,我们可能得出结论,他并不高度看重睡眠。这可以从著名的引语如*“坟墓里有的是睡觉的时间”或”睡着的狐狸抓不到家禽”中判断。这种态度类似于今天那些准备实践多相睡眠的人。这也是我们对睡眠生物功能知之甚少的时代高成就者的常见特征。然而,富兰克林更著名的是说:“早睡早起使人健康、富有、明智。”* 由此我们可能得出结论,如果他想少睡,他的公式是早起,而不是把睡眠撕成碎片。此外,对于一个不太看重睡眠的高成就者,退休小睡可能感觉是懒惰或软弱的主要标志。这种污名在西方文化中持续至今,小睡常被视为懒汉的习惯。最后,作为 DST(夏令时)的倡导者,富兰克林会说:“人们大多靠烛光生活、在阳光下睡觉,这是愚蠢和浪费的。” 他绝对不是多相睡眠者。
温斯顿·丘吉尔(Winston Churchill)
我们对 Winston Churchill 的睡眠习惯了解颇多。作为战时首相,他的日常作息被助手密切关注。丘吉尔可以工作到深夜让部长们精疲力竭,但他也会规律地在下午进行扎实的 1-2 小时小睡。因此,他是一位经典的双相睡眠者。在 Chartwell 的家中,他的作息相当规律:8 点醒来,在床上度过早晨读报、口述信件等,在茶点时间长时间小睡,工作到凌晨 3 点。他平均每天睡 5-6 小时。这些话被归于丘吉尔本人:“你必须在午餐和晚餐之间睡觉,不能半途而废。脱掉衣服上床。我一直这样做。不要以为你白天睡觉就干得少。那是没有想象力的人持有的愚蠢观念。你能完成更多。你一天当两天用——嗯,至少一个半。” 丘吉尔训练有素的双相习惯使他成为最有活力的战时领袖之一。幽默地讲,F. D. 罗斯福的助手注意到,丘吉尔来访后,美国总统如此疲惫,需要连续 3 天每天睡 10 小时来恢复。
托马斯·阿尔瓦·爱迪生(Thomas Alva Edison)
托马斯·阿尔瓦·爱迪生与睡眠有着爱恨交织的关系。睡眠研究者指责他剥夺了现代人口 1-2 小时的睡眠。工作狂喜欢引用他对睡眠的蔑视。多相睡眠支持者声称他是多相睡眠者。确实,爱迪生对睡眠的蔑视有充分记录,但这只能归因于他的无知。在他那个时代,对睡眠生物作用的了解甚少。他错误地相信,如同食物一样,人类如果有机会总会睡得比必要的多。作为天生的短睡眠者,他相信长睡眠是懒惰的标志:“大多数人过量进食 100%,过量睡眠 100%,因为他们喜欢。那额外的 100% 让他们不健康和低效。每晚睡 8 或 10 小时的人从不完全睡着也从不完全清醒——他们在 24 小时里只有不同程度的打盹。” 在一次并行的无知闪现中,爱迪生看不到体育锻炼的多少价值。他的冬季住所配备了一个最早的现代游泳池,但爱迪生从未使用它。他只是不认同运动和睡眠是心理和身体健康良好投资这一现代观点。他的同事注意到,爱迪生实际睡得比他愿意承认的多得多。显然,他把少睡当作荣誉徽章。他经常打盹,他的小睡床至今保存在爱迪生博物馆。然而我找不到任何可信证据表明,爱迪生的小睡遵循”困了就睡”以外的任何规则,而这通常匹配双相模式,或至少符合 PureDoxyk 定律。我能找到的关于爱迪生睡眠的最可靠信息,是他接近 40 岁时短暂保留的一本日记。从这本日记我们可以了解很多他的睡眠习惯。他似乎相当痴迷于获得良好的夜间睡眠,因为他的日子常常以关于睡眠质量的笔记开始。像我们大多数人一样,他睡得越好,似乎越快乐。这与多相支持者的主张恰恰相反。爱迪生宁愿最大化他能使用神清气爽头脑的小时,而非最大化清醒小时。而从睡眠生理学、心理卫生和生产力的角度来看,这恰恰是最合理的。
尼古拉·特斯拉(Nikola Tesla)
在爱迪生门下短暂效力后,尼古拉·特斯拉成为前导师的激烈对手。我们都听说过”电流之战”,但爱迪生和特斯拉在另一个战场交锋:他们试图在少睡上互相超越。特斯拉注意到爱迪生睡得比他愿意让别人相信的多得多。这给特斯拉和爱迪生自己关于实际睡眠量的报告注入了一丝吹嘘人格。我打赌同样的机制使今天的博主常常夸耀多相适应。特斯拉确实能通宵工作,但在其壮举后常常崩溃睡一整天。他表现出狂躁创造力的经典迹象,可能被短暂的恢复性小睡或漫长的恢复性睡眠打断。除此之外,特斯拉不过是个短睡眠者。他太忙于自己的追求,从未想过任何类似严格多相作息的东西。那会是对他张扬人格的束缚。
总而言之,整个多相天才名单似乎缺乏任何可信证据。因此,它可能是那些想给自己的一天增加清醒时间的人集体一厢情愿的产物。
多相睡眠的可持续性(Sustainability of polyphasic sleep)
多相睡眠的主要问题源于:
- 在睡眠禁区难以入睡,
- 难以从深的主观夜间睡眠中醒来,
- 生物 clock 的可训练性有其极限,使得绕过问题 (1) 和 (2) 成为不可能。
多相睡眠的时钟与沙漏模型(Clock and Hourglass model of polyphasic sleep)
如果我们使用时钟与沙漏隐喻,可以用简单的术语解释为什么对多相睡眠的适应永远不会发生:
- 早上,如果你神清气爽休息良好,你的催眠药水(即昼夜节律困倦)被清除,你的能量沙漏充满精神能量(即你的内稳态困倦被清除)。你不太可能在早上入睡。在那个时候试图小睡是浪费时间,你会浪费时间准备小睡,浪费时间试图入睡。
- 下午,在午睡时间,有一个由生物钟控制的警觉性低谷。当你的能量沙漏越来越空时,你可能能小睡。这没问题,你的小睡会很短,因为夜间催眠药水不在。
- 傍晚,你的催眠药水仍不在。如果你下午小睡了,你的能量沙漏几乎充满能量。如果你试图再小睡,你会盯着天花板,再次浪费时间。
- 夜晚,你的催眠药水被释放。小睡应该容易,但如果你入睡,你不会醒来——不会自然醒来。你需要闹钟。你可能设法快速重新填充沙漏,但催眠药水会让你昏沉疲倦。你可能需要双重闹钟或响亮的闹钟,或你妈妈的帮助(如果她曾同意这个多相实验)。你会战斗和挣扎,你永远不会自然醒来。只要催眠药水在起作用就不会,当你的昼夜节律系统说这是半夜时就不会。
如果你决定多相睡眠,你必须使用闹钟,否则你不会在夜晚醒来。一旦你使用闹钟,你就会睡眠剥夺,这会让你的能量沙漏方便地被排空。空的沙漏确实会让小睡更容易,但决定你脑力的是沙漏。沙漏空了,你不过是个头脑空空的行尸走肉。要以等间隔产生小睡,你必须杀死困倦的 24 小时昼夜节律组分,必须杀死你的生物钟并阻止催眠药水的释放。这是不可能的。催眠药水会每 24 小时释放一次,让你困倦;无论你怎么挣扎。最短的自然夜间睡眠很少低于 3 小时。许多双相睡眠者可以靠 4 小时过得很好。然而,大多数青少年可能需要 7 或 8 小时夜间睡眠才能最佳运作。在健康的睡眠中,白天小睡要么不可能要么非常短。如果你用 SleepChart 免费版追踪睡眠,你可以自己看到这一切。你会看到小睡如何倾向于在夜间聚集(对你可能是中午)。这正是多相大师 Stampi 博士在单人水手身上观察到的。记住,要让这个图景成立,你应避免闹钟,而这在多相睡眠中自然不可能。然而即便在强制作息下,你会看到小睡在夜间更长更频繁的规律模式(每次你放松纪律、睡过头等)。白天小睡会更短,尤其是在主观傍晚时段(对你可能是午夜)。
生物钟训练的极限(The limits of the body clock training)
我一次又一次听到,所有生物学推理都无关紧要,因为身体总能适应训练和压力,且科学尚未研究成功的多相睡眠者。以下是基于时钟沙漏模型的回复:
- 生物钟由基因控制,我们不知道能显著影响生物钟周期的药理因素。多相睡眠需要把生物钟周期缩短六倍!另一种可能是完全移除生物钟,让精神能量沙漏来支配睡眠周期。
- 生物钟相位可以用光、活动、褪黑素和其他因素移动,但释放催眠药水的周期长度难以控制。药物可以减少催眠药水的影响,但这应避免,因为它影响睡眠阶段周期。用磁盘和内存隐喻来说,并非你所有 PC 数据都能写入硬盘并被碎片整理。
- 能量沙漏被排空的速度可以受药物(如咖啡因)影响;然而,更快的沙漏会产生更多睡眠(而非更少),而更慢的沙漏会使多次小睡更不可能。
- 多相睡眠在实验室条件下,如果用闹钟打断自然睡眠是可能的。驯化的自由运行多相睡眠在健康个体中不可能。
- 科学尚未研究成功适应的、具有自然多相节律的多相睡眠者,因为他们不存在(尽管有许多该头衔的声称者)。Stampi 博士的实验不符合条件,因为它们总是涉及闹钟。
结论(Conclusion)
健康的生物钟运行 24 小时周期。这个周期会让你在主观夜晚(也可能对你是中午)困倦。这就是为什么没有闹钟你无法在主观夜晚从小睡中醒来。闹钟不健康,它们阻止睡眠履行其功能。选择是你的:(1) 多相睡眠,或 (2) 自然睡眠并让你的大脑发展其全部智力潜能。如果你仍不信服,请阅读 Uberman 睡眠发明者的这条消息。
多相睡眠中的咖啡因(Caffeine in polyphasic sleep)
多相睡眠者相信,避免咖啡因可能有助于适应。由于咖啡因消除相对缓慢及其对腺苷受体的影响(抵消内稳态困倦),在小睡前 5-7 小时以上摄入咖啡因据说会使小睡更困难(除非摄入就在小睡前)。
确实,避免咖啡因会使多次小睡更容易。然而,它不会解决在主观夜晚时段醒来的昏沉问题。多相睡眠的问题在于昼夜节律周期的不对称(只受咖啡因轻微影响)以及内稳态困倦的缓慢累积。即便完全戒除咖啡因,也不会产生足够的内稳态困倦来确保在所有期望时段小睡。反过来,服用强效腺苷激动剂更可能导致类似发作性睡病的睡眠模式,而非期望的 Uberman 睡眠。这恰恰与多相信徒少睡的目标背道而驰。多相睡眠模式本质上不稳定,咖啡因水平的变化对这一事实没有任何影响。
至于正常的健康睡眠(多相睡眠不是),戒除咖啡因并非必要,但所有咖啡因饮料应最佳地只在醒来后的头两小时内饮用。
多相睡眠突变体(Polyphasic sleep mutants)
一些多相睡眠信徒想知道,博客上罕见的成功报告是否由于某种使这些个体更可能成功的突变。这在理论上是可能的,但极不可能。要让”突变体理论”成立,我们需要一种产生没有昼夜节律组分睡眠的突变。这样的突变实际上已知,并导致源于持续睡眠剥夺的严重残障。受这种突变影响的人永远不会是正常的睡眠者(像多相睡眠信徒)。另一种突变可能允许 homeostatically 产生类似昼夜节律低谷的状态,这种状态周期性出现在所有脊椎动物的大脑中。这就像想象一种允许人以 25 克份量排便的突变,或允许异步自愿蠕动的突变,或用两个独立调节的眨眼机制取代眨眼反射的突变,或交替 3:6:3:9:3:6 间隔比的持续切分心律,或心房单独收缩,或心室单独复极化等,或可以被轮班工作驯化、每 9 天出血一次的月经周期。最接近可以匹配多相睡眠可能由突变启用这一假设的障碍是发作性睡病,患者确实在一天中多次打盹。然而,这是一种不扁平化昼夜节律功能的内稳态障碍。因此,发作性睡病患者睡得比健康人多,而不是少。1996 年,研究者能够通过玩弄光照暴露使西伯利亚仓鼠无节律(Ruby 等,199656)。然而,他们的生物钟仍在运行周期并对光诱导的相位移动做出响应,只是运动活动节律变得解耦。我们知道,人类的无节律性会因睡眠无法在没有昼夜节律组分的情况下履行其功能而导致严重残障。此外,很难把人类的遗传学与生活在寒冷气候、在地下洞穴中度过延长黑暗期的动物相比较。类似的基因”适应”多相睡眠的可能性,可能与人类冬季头发变白的概率相当。
最后,我怎么分辨一个多相突变体?他/她很可能从出生起就多相。尽管 PureDoxyk 从未真正像 Uberman 那样多相,她的睡眠模式一直有些不规律地多相,这正是使她的案例可信的原因。在遗传学上,双相睡眠与驯化良好的超日节律多相睡眠相去甚远。甚至婴儿都几乎不是超日节律的(见:婴儿睡眠)。换言之,当一个其他方面健康的人突然声称多相适应时,我只能严重怀疑。
多相睡眠博客(Polyphasic sleep blogs)
由于我无法进行自己的多相实验或鼓励他人多相睡眠,我通过阅读网上的多相博客收集了大量关于 Uberman 概念的洞见。这样的博客有几十个,它们提供了相当有趣的阅读。除了与困倦或昏沉的持续斗争,这些博客也流露出对健康睡眠原则的缺乏理解和对睡眠重要性的总体漠视。这里有一个有代表性的引用:“如果一个人活 75 年,他们会有 25 年是无意识的。那是我直到现在的整个生命,被完全抹去、未被使用。家庭、学校、工作、写作、你们所有人,没有发生过。那就是每天睡 8 小时的代价。所以我把我睡眠削减到 2 小时,试图从短暂生命中榨取一切。” 对于无法欣赏睡眠作用的人来说,这句话听起来可能不像应该的那样令人愤慨。然而,由于我们大多数人欣赏工作的价值,这在我看来大致如下:“如果一个人活 75 年,他们会有 25 年在工作。那是我生命的大部分,被完全抹去、未被使用。家庭、学校、睡眠、写作、你们所有人,我都没时间。那就是每天工作 8 小时的代价。”。
关于幽默的生物学基础的一种理论说,它产生于对其他个体的优越感。据说,那些能够检测同类无知的人,通过幸福感和快乐感强化其发现。因此,看到他人做愚蠢的事是有趣的(只要希望在这个过程中没有人受伤)。据推测,取笑愚蠢者的演化机制,帮助人类在有书面记录之前很久,通过世代保存智慧。
在这一背景下,如果你理解暗示多相作息驯化不可能性的睡眠控制机制,你可能会发现研究多相睡眠者的博客极其有趣,实际上是搞笑的。到处都是线索和危险信号,博主们却不断撞墙。幸运的是,这些个体通常在几周痛苦后看到光明。我们应该希望在这个过程中没有人受伤,例如因睡眠剥夺状态下驾驶。
所有博客似乎大致经历相似阶段。它们以对 Uberman 睡眠改变生活的潜力的青春亢奋开始,整个概念带有一种邪教光环。它与 Aum、大卫教派、太阳圣殿教或人民圣殿教的工作伦理和自我强加或外部强加的睡眠剥夺平行。这种修道式的吸引力因野心勃勃的采纳者常常把各种形式的节食作为其”改革”的一部分而加强。与”多相实验”相关的希望很多,通常围绕能做更多、体验”增加的能量”。希望被许多志愿者首先难以获得令人神清气爽睡眠这一事实放大。然后斗争开始,点缀着对”暂时适应阶段”的希望性引用。一切都始于昏沉、醒来问题和睡过头。疲倦累积,词频分析显示”tired(疲倦)“是这些博客中最常用的词之一(连同”I”和”nap”)。然而,快乐的”polynapper”通常能凭借热情加上额外时间和三倍能量执行生活重大改变带来的兴奋度过初始阶段。然后实验的负面方面开始显现,包括无法克服的困倦、穿过精心设计的闹钟系统睡过头、体温调节问题、负面躯体症状、因反复睡过头的自责等。睡过头反复在主观夜晚发生,而小睡问题在主观白天发生。然而”polynapper”对注意到这种规律很迟钝。他们为获得额外清醒时间而高兴,然而他们非但没有高效利用,反而拼命寻找任何消磨时间的东西来”熬过雾”。他们在错误时间徒劳地试图入睡上浪费宝贵时间。当事与愿违时,他们开始试验睡眠作息的各种变体:更多小睡、更少小睡、更长小睡、更短小睡、“伪小睡”、固定作息或”弹性小睡”等。由于这些通常徒劳,“核心睡眠”或”恢复性睡眠”的概念开始被考虑。一些实验者决定”倾听身体”。有了”核心睡眠”和对自身身体节律的一些关注,实验者向双相睡眠的变体漂移,并可能逐渐接近合理的睡眠作息。然而,不理解睡眠调节双组分模型的基础,很难弄清楚自己的最佳睡眠时机。困难被两个因素加剧:
- 相信多相睡眠模型会奏效,以及
- 昼夜节律中失去同步。
至于后者,驯化良好的自由睡眠相对容易理解。然而,一旦强相位移动刺激被引入系统,尤其是异步应用或更糟的不规律模式,整个睡眠控制系统变得混乱,基本上不可预测。换言之,即便是经验丰富的时间生物学家也可能发现难以解释睡眠区块时机与警觉性之间的相关性。如果不幸的实验者看不到双相的光,他就开始理论化自己无法坚持作息的原因。这些可能是坏食物、坏激素、缺乏自律、跳过小睡、额外小睡、工作麻烦、朋友、睡眠过多、REM 过多、REM 过少、“第 4 阶段 REM”过少(原文如此!)等。理论家们说得好像人能轻易猜出”组胺水平”或小睡中”第 3 阶段睡眠”的持续时间(无需验血或脑电图)。在健康的自由运行个体中,3-5 分钟内入睡应是轻而易举的事,然而多相睡眠者却在昼夜节律高峰时不断与无法足够快入睡作斗争。同样艰难的是,他们在昼夜节律低谷时从小睡中醒来。难怪睡过头继续,与困倦的斗争开始产生代价。最终,博主通常把实验推迟到”更好的时候”(圣诞后、假期后、疯狂时期后等)。有时博客只是没有结论地突然结束。“polynapper”很少承认失败,或总结多相睡眠不可行。少数人不诚实地声称成功适应了睡眠作息,然后转向其他主题的博客。
这些年轻男子往往渴望生活、渴望经验、渴望成就,无法适应晚上 10 点 - 早上 5 点的睡眠作息,叛逆并准备寻求最大生产力的新方式。这些大多是高尚的特征,但与无知混合时,可能导致糟糕的健康、糟糕的决策、糟糕的脑力表现和社会摩擦。这些人格类型也面临更高的早逝风险。多相睡眠可能也有其贡献:“我刚从加拿大一路多相驾驶过来。” 多相睡眠只有一个主要好处:多相博主对我们理解睡眠做出了贡献。没有研究者能在伦理上让这么多个体承受多相作息的精神折磨。在本文:Polyphasic sleep: Myths and Facts : Comic Relief 中,我汇编了多相博客中最有趣的引用清单。这些说明了实验的各个阶段,特别关注睡过头和警觉性。自然,这个清单非常有选择性且脱离上下文。博主常常声称他们感觉很好、方法奏效、他们计划无限期继续。然而与热情交织的是危险信号,令人惊讶地不断被忽视。几个博客甚至大声宣称巨大成功。我不会引用或链接这些,因为我觉得它们相当不诚实,或带有隐藏议程。这些会稀释真相并炒作一种潜在有害的生活方式。
多相迷思(Polyphasic myths)
阅读多相睡眠博客时,可以识别出一些在博客间像坏性病一样不断传播的迷思:
- 错误! 大多数动物多相睡眠,因此人类也必然如此。
- 错误! 适应期艰难但会在某个时刻结束。
- 错误! 多相睡眠节省时间。
- 错误! 多相小睡只含 REM。
- 错误! 多相睡眠让你更警觉。
- 错误! 多相睡眠让你更高效。
- 错误! 多相睡眠作息让你减肥。由于睡眠剥夺对食欲激素的影响,很可能发生相反的情况。
- 错误! 多相睡眠是健康的。
- 错误! 长小睡对你有害。
- 错误! 即便不睡眠剥夺,多次小睡也比一次小睡好。
- 错误! Claudio Stampi 向所有人推荐多相睡眠。
- 错误! 多相睡眠最大化个体获得的 REM 量。
- 错误! 历史上许多天才多相睡眠。
要阅读多相博客的一些有趣摘录,见:Polyphasic sleep: Myths and Facts : Excerpts from polyphasic sleep blogs。
影响睡眠的因素(Factors that affect sleep)
压力(Stress)
压力是睡眠杀手。与压力相关的激素,如肾上腺素、ACTH、皮质醇等,会增加警觉性并降低睡眠倾向。在压力条件下更难启动睡眠,夜间醒来更可能发生,睡眠结构也可能改变,睡眠可能变得更短、更不令人神清气爽。此外,睡眠剥夺会放大压力情境的影响。压力和糟糕的睡眠联手,使工业化国家相当多人的生活变得悲惨。那些困倦且有压力的人更不可能实现目标,这只会加重痛苦。
除了对睡眠的影响,压力还可能对创造性工作、学习、问题解决等产生负面影响。在最坏的情况下,压力几乎能让你的大脑萎缩!持续的压力和升高的皮质酮水平已被证明会降低大鼠大脑中的 BDNF,导致海马体(主要的记忆交换机)萎缩。压力可以降低你的智商,不仅是降低一天,而且在较小程度上降低一生!
压力根植于我们的情绪大脑。情绪非常难以控制,常常决定一个人的成功或失败机会。诸如愤怒等负面情绪是适得其反的,对个人成长贡献甚微。诸如剂量恰当的激情等正面情绪,帮助人克服任何赛道上必然遇到的障碍。情绪也是可传递的,并倾向于在社交群体中放大。愤怒滋生愤怒,热诚滋生热诚。在合适的情境下,你努力产生正面情绪,将向你互动的社交圈发送正面涟漪。学会利用正面情绪并绕开负面情绪。邀请所有帮助你执行宏大计划的正面情绪,让自己爱上你的工作、人和世界,远离负面情绪的来源。
情绪的力量来自这样一个事实:它们被接入低层脑结构,无法轻易地被由前额叶皮层主导的理性思考所控制。一个愤怒的人可以命令大脑冷静下来,但他无法瞬间降低已经释放到血液中的肾上腺素水平。一个吸毒者可以理性地决定戒毒,但当渴望的身体效应袭击他的系统时,他的理性大脑常常无力。
由于难以对抗自己对压力的反应,应对压力的最佳方式之一是逃离它。压力对你的福祉如此重要,如果可能,应考虑换工作、换朋友、换住所或换生活方式!没有这些,即便最好的抗压建议也可能无效。有些人天生易受压力影响,可能发现无法过没有压力的生活,另一些人则可能在压力中茁壮成长(在一定程度上)。本文甚至无法触及这个麻烦冰山的一角。如果你饱受糟糕睡眠和压力之苦,应对压力可能是你的头等大事。关于压力的书籍和博客汗牛充栋,再列一份清单似乎多余。然而,我想按我认为可能有帮助的程度(尤其是从睡眠和创造性工作的角度)做一份排序选择。我相信按信息力而非治愈力排序很重要。例如,良好的健康可能是对抗压力最重要的因素,然而你可能已经在为之努力。另一方面,较少人意识到痛苦在治愈烦恼中的有效性!我不是压力管理专家,而且我有幸拥有相当强的耐压能力,这使我难以列出全面的清单。如果你认为我可以添加值得向他人推荐的内容,请告诉我。以下是我的清单:
- 运动:如果你施加运动的痛苦,你的其他痛苦可能会从新的视角被看待。试试马拉松或冬泳。很少有东西能像痛苦一样带来如此多的解脱和快乐!运动帮助你生长大脑——你最好的抗压盟友。另一方面,睡眠剥夺和压力使大脑萎缩,让人更难在生活中挣扎。
- 消除打断:只要可能,在一天中创建你可以专注于创造性工作的保护区。如果可能,整个早晨都应受到保护以保持头脑清醒。在那个时段,电子邮件、电话和会议都被禁止。如果保护区紧接在压力期之后,你可能永远无法回到最佳心理状态。偶尔进行创造性的假期,期间无人能联系到你。如果这太多,试一次,也许你会从新的角度看整个生活。
- 测量一天:与其测量你与目标的距离,不如测量你在给定的一天完成了多少。如果你的目标足够雄心勃勃,大多数时候它们会在远方的地平线上。眼睛盯着地平线,你对进度没有良好感觉。如果你专注于一天的工作,你会更好地感觉到积极的运动。不要等待大奖杯,让小小的好事成为自己的奖赏。一旦你在小事上达到更高的满意度,你的大脑可能以更高挡位运作,也许把你带到原始目标之外。
- 简化:简单的生活更有趣。简单是逃离激烈竞争的绝佳方式。一些想法:骑自行车而非开车,放弃一些电子必备小玩意,尝试无截止日期地生活,少做承诺(包括对自己),关掉手机,进行创造性假期(一份工作、一个目标、无干扰)等。如果你同时做太多项目,尝试在开始另一个之前完成一个。学会说”不”或避免不得不说”不”的情境,学会委派。如果你信息或邮件过载,把它们全部倒入 SuperMemo 并排序。
- 创造性工作:我相信很少有东西像为好的事业进行的创造性工作那样令人满足。如果你能为一个主要的创造性目标牺牲次要的东西,每次有机会都这样做。从长远看,头脑的生产力是幸福健康生活的最佳公式之一。
- 慢下来:如果你总是觉得没时间做事,你可能给自己盘子装得太多。这会放大你的压力水平,给你推迟睡眠的借口。快速高效是好的,但这不能超过某个生物学极限,在那里你用今天的次要成就透支你的生活。
- 排序:在长远看堆积压力的好方法之一是拖延或做有趣的事而忽视其他。你需要始终诚实地排序并按优先级一丝不苟地执行。清单底部总有一长串需要被忽略的。忽略过载、专注于最高优先级工作是每个人都需要学习的。有效排序的方式之一是使用按优先级 = 价值/时间排序的任务清单(tasklist)。把任务倒入任务清单既无压力(你总是希望某个时候执行)又高效(你总是从清单顶部工作)。
- 睡眠:尽管我们把睡眠当作目标,睡眠也是清除压力的好方法。然而,这把武器只有在你自由运行睡眠时才有效,因为压力本身可能导致相位延迟。这意味着你可能需要稍微推迟睡眠并起得晚一点。如果你需要早起,睡眠不太可能是你的盟友。如果你陷入破坏一天的不安情境,你总可以寄望于一些扎实的运动加上扎实的睡眠。坚持人类生物学会为你工作的舒适想法。压力情境在压力累积时似乎更有压力。压力倍增压力,良好的睡眠给你今天担忧一个新的、更诚实的视角。
- 友善:如果你有正面的东西要对某人说,就说!帮助他人带着微笑度过一天。如果你心里有负面的东西,思考表达它是否必要。小事不值得为之争斗。当你心情不好时,记住,即便你强迫自己对他人友善一点,你会发现友善是有感染力的,你会得到一些回报,一天会感觉更阳光。如果这个建议不起作用,也许你在错误的环境?
- 家庭:对许多人来说,家庭是压力的主要来源,但它也可以是压力缓解的最佳形式。如果你即将组建家庭单位,记住,长期的无压力生活应是你主要目标之一。如果你的关系是一连串担忧而没有隧道尽头的光,也许它不是你最好的长期投资?
- 自然:可悲的是,我们中许多人失去了从自然世界之美汲取力量的能力。对于一个压力大的工作狂,去森林或山里的旅行感觉像是浪费时间,只会放大压力。然而,对自然的爱在每个人身上无意识地休眠,它可以被重新唤醒,可以重新学习。一旦恢复,与自然的接触是最好的压力缓解剂之一。毋庸多说,自然保护区应是手机禁区!
- 知足:提升一个人抗压能力的最佳方式之一,是致力于整体知足感。研究表明,知足主要基于自尊、关联性(与他人的关系)、自主性(掌控感)和能力(生产力)。在一定程度上,所有这些因素都可以被影响。知足让人更容易忽略小烦恼、看清大局和大目标。
- 避免冲突:只为改变人生的大事业而战。这个星球上太多时间花在了为或反对小事和/或次要原则而战。大量冲突可以通过忽略它并专注于更重要的事来解决。
- 避开坏人:压力的主要来源之一是他人。一些是好人,因工作、职责、好意、无知等带来压力。另一些纯粹是坏人,最好避开。在选择配偶、工作、健身房、朋友等时,确保远离那些有 spoiling 你一天天赋的人。这些选择将决定你的压力水平、睡眠能力和专注重要事情的能力。
- 提前计划:当你带着一天的目标醒来,你的心思从压力性干扰中移开,生产力飙升。定期回顾你的目标清单以重新聚焦心思。你需要为工作、家庭、运动等分别设立目标,它们都有助于每个每天顺利高效地航行。
- 时间管理:扎实的时间管理对生产力至关重要。低生产力滋生低生产力,一旦效率下降,就会对你的动机产生负面影响,进一步削弱你的进展。就烦人的压力而言,很少有比堆积的未完成事项清单更糟的了。那些烦人的清单最好用时间管理(完成事情)和排序(去除无法完成的事)来解决。考虑 SuperMemo 中的 Plan 进行时间管理,Tasklists 无压力地砍掉待办清单底部。
更多内容,见这篇不朽文本:The Medical Basis of Stress, Depression, Anxiety and Drug Use!。
正面压力(Positive stress)
一定程度的压力也可以是正面的力量。一些形式的压力是极好的能量剂。我相信,在为好睡眠、好学习和好创造力优化一天时,考虑压力的时机是有道理的。睡前的压力会对睡眠产生负面影响,创造性工作或学习前的压力会对脑力工作的结果产生负面影响。绝大多数人对压力的时机没有多少影响,压力似乎渗透我们所有的生活。然而,如果你属于幸运的少数,可以决定何时打开信件、读邮件、打一个困难的电话、安排艰难的会议、处理压力性任务,你应该尝试让压力为你所用。我相信,处理压力性情境的最佳且唯一正确的时机是午睡前或午睡时。这一时机节省了最具创造性的早晨时段,并在夜间睡眠前提供了足够的时间缓冲。此外,它帮助你航行通过一天中生产力较低的部分,包括中午警觉性低谷(如果你负担不起小睡)。如果你是小睡者,在压力时段后加运动可以相当高效地冲走压力的影响。一剂压力实际上可以提高运动的效率,如果运动不足以消除压力的影响,你只会牺牲日常睡眠周期中较次要的组成部分:中午小睡。
酒精(Alcohol)
酒精是创造型个人的主要敌人!过量时它对大脑高度有毒!即便是小剂量也会降低 REM 睡眠的质量和密度。酒精还抑制深睡眠,产生睡眠碎片化,并放松上气道肌肉,这会加重打鼾和阻塞性睡眠呼吸暂停的严重程度。除了对睡眠的负面影响,酒精降低认知能力、抑制记忆编码,在创造性努力时期应特别避免!
另一方面,大量研究表明,小剂量的酒精可能有益健康。实际上,每天一杯酒可能是预防动脉硬化、心脏病发作和脑血管疾病的最简单的已知方法。有报告称适度饮用啤酒,甚至一般的酒精,可能降低阿尔茨海默病的发病率(Breteler 等,200266)(啤酒肚或啤酒罐中的铝会有相反效果)。少量酒精可以改善血脂谱,而相比之下,过量饮酒与高血压相关。一些医生推荐每天极少量饮酒(不超过一杯)。
对于一个高度创造性的人来说,酒精构成了健康与大脑的两难。它肯定应在睡前 3-5 小时避免,在智力工作之前如果中间没有睡眠也应完全避免。这将只留下在傍晚早些时候(假设你之后不做任何脑力工作)或午睡时间(假设这是你从脑力努力中休息小睡的时间)非常适度饮酒的空间。
运动已知会减少饮酒,可能是通过对奖励中心的影响。然而,它也应提供一些保护,抵御酒精对大脑的毒性作用。运动加速循环并加速酒精转化为乙酰辅酶 A(acetyl-CoA),然后可以快速用作能源。这防止了乙醛(乙醇最毒的代谢物)的累积。乙醛部分导致宿醉,并可能有致癌性。运动还帮助降低血液甘油三酯水平,这可能因慢性饮酒而升高。规律的适度饮酒改善中和酒精的代谢机制。另一方面,狂饮等同于摧毁自己的大脑。如果你曾出现言语含糊或经历宿醉,你就知道你的大脑发生了不好的事!你越年轻,预期的损害越大!
你绝不应在睡前饮酒。酒精被快速代谢,会产生乙醛反弹效应,大大增加夜间醒来的机会。这种效应让酗酒者夜间失眠,剥夺他们的 REM 睡眠,可能实际上负责震颤谵妄(delirium tremens)(也许还有Korsakoff 精神病)。即便是适度量的酒精也会对睡眠质量有明显影响!确保酒精在你夜间睡眠前离开你的系统! 睡前一本书可能和一杯葡萄酒一样有效!
你绝不应在创造性工作或学习前饮酒。哪怕一口啤酒都可能影响你的表现。一些 SuperMemo 用户声称他们享受在晚上学习时适度饮酒。如果晚上学习的功能是娱乐和放松,对学习效果本身没有太大期望,这是可以理解的。我不确定这是否会恶化或缓解记忆过载对海马体及相邻网络的影响。然而,我确实知道,由于傍晚下降的昼夜节律斜坡和酒精本身,记忆效应会大大降低。
你可能考虑的一些饮酒规则:
- 夜间睡眠前血液中的酒精会严重降低睡眠质量
- 酒精抑制创造力
- 酒精使学习无效
- 宿醉对大脑有害,表明过量
- 言语含糊或平衡问题表明过量
- 如果你违反过量规则(如上),暂停酒精一个月,向自己证明你没有走向成瘾
- 试着每杯酒至少搭配一小时运动来保护大脑
更多阅读:
咖啡因(Caffeine)
咖啡因是用于对抗困倦的头号药物!90% 的美国人以某种形式使用它。它存在于咖啡和可乐中,在茶和巧克力中也有少量。它具有成瘾性,通过类似于安非他命和可卡因的通道起作用,还影响奖励中心(包括伏隔核(nucleus accumbens)(Goldberg 等,200267))。由于咖啡因通过阻断腺苷受体对中枢神经系统产生深远影响,它被广泛用于对抗困倦。然而,大多数人很少意识到,它在对抗与腺苷相关的内稳态困倦组分时效果良好,但在克服昼夜节律困倦时相当低效!此外,用于对抗后者时,它实际上可能相当不健康!如果你滥用咖啡因,或在生物钟告诉你睡觉时使用它,你只会体验到那些给咖啡因带来所有坏名声的症状。这些包括:心律失常、易怒、压倒性的疲倦、抑郁、流产风险增加,以及典型的咖啡滥用者的”胃部不适”。难怪流行迷思说咖啡对健康有害,可能导致心脏病。
令人惊讶的是,关于咖啡因健康影响的研究似乎并未证实其有害性。考虑到咖啡的制造方式,其致癌效应微不足道似乎令人惊讶。一些出版物甚至指出咖啡对健康和长寿有正面影响。如果研究看起来矛盾,可能来自这样一个事实:一些人在”正确的时间”喝咖啡,而另一些人试图代偿睡眠剥夺或掩盖昼夜节律困倦。咖啡与心脏疾病之间的联系微弱,可能取决于个体代谢咖啡因的基因能力,并可能归因于以过量剂量或错误时机剂量形式的咖啡因滥用。一些研究甚至发现 3-5 杯咖啡对寿命最佳。同一研究被批评未能注意到,咖啡在更富裕、有利于长寿的家庭中更受欢迎。也有关于咖啡因对记忆有正面影响的报告。咖啡因增加海马体中 BDNF 的水平,可能促进神经发生,被发现对阿尔茨海默病有适度预防作用。那么你可以假设,咖啡因在较小的 200-400 毫克/天(相当于 2-4 杯咖啡)时相当无害。注意 50% 的美国人摄入更多。要让咖啡因无害,必须在正确的时间摄入!
咖啡因倾向于把许多人驱动到恶性循环:你喝它,肾上腺素(epinephrine)激增,你感觉更有活力,多巴胺(dopamine)激增,你感觉更好,你觉得自己能熬夜,你睡得更少,第二天你更困,所以你需要更多咖啡因,由于下调你每杯得到的提升更少,你纳闷为什么这次不起作用,你增加剂量,恶性成瘾循环继续。咖啡饮用者偶尔会经历偏头痛样头痛,这些由咖啡因供应较少的日子腺苷受体活动增加引起,导致大脑血管扩张。血管扩张或感觉神经元上嘌呤受体的激活产生头痛。半杯正常剂量的咖啡因应有帮助。结论:如果你想戒除咖啡,不要突然完全戒断,留出几天让身体逐渐对抗成瘾。对咖啡因的理性方法是:把它用作昼夜节律增强剂!尽管我总是建议避免用物质调节睡眠,但你可以用咖啡因加速从睡眠到完全心理警觉的过渡。早晨的小剂量会让你警觉性斜率上升,规律摄入会产生温和成瘾,应有助于你在傍晚入睡。这种方法对你的健康、睡眠结构应基本中性,对警觉性(至少在一天的很早时段)正面。绝不要用咖啡因掩盖睡眠不足! 当前关于咖啡因的知识支持在健康个体早晨喝一杯咖啡的建议。由于黑咖啡可能刺激胃黏膜,咖啡应用牛奶或奶油饮用。在不与失眠斗争的规律小睡者中,昼夜节律仍应允许从下午小睡醒来后立即喝咖啡。
作为唤醒药物,咖啡因可能诱发失眠。这就是为什么它绝不应在睡前 6-7 小时之后摄入。咖啡因半衰期对健康个体约 6 小时,但个体间可能差异巨大!摄入过晚,咖啡因会抑制 REM 睡眠,损害睡眠质量及其对记忆的影响。同时,在一天早些时候规律摄入时,它实际上可能产生温和的戒断效应并促进睡眠。如果足够早服用,它对睡眠结构的影响也应轻微,然而,即便是早晨摄入也会减少夜间深睡眠(Dijk 等,199568)。咖啡因不能作为对抗睡眠剥夺的武器,只有充足的夜间睡眠才能扮演这个角色。咖啡因也不应用于对抗昼夜节律睡眠组分。正如全文所论证的,昼夜节律最好留给它自己运行!
一些睡眠专家推荐小睡前喝咖啡以缩短其时长和/或确保从第 2 阶段 NREM 醒来。如果你需要匆忙快速恢复性小睡(例如困倦的驾驶员),这可能没问题。然而,自由运行周期中的最佳小睡自然不会超过 30 分钟,尤其是在压力条件下。如果小睡被一杯咖啡人为缩短,小睡的效果可能短暂。
喝咖啡唯一的好时间是早晨!(或在习惯性小睡者小睡之后)
绝不要喝咖啡来克服昼夜节律困倦!
Stickgold 博士说:“极有可能,绝大多数早晨喝咖啡的人,是有意识地或无意识地,为了纠正因睡眠量或质量不足导致的困倦。” 不一定如此。即便在一夜好眠、没有睡眠剥夺之后,咖啡也能提升大脑的创造力。然而,它很可能以注意力为代价,可能有助于创造性问题解决,但也可能减少你早晨讲座的注意力或放大压力情境的影响。最终,你需要做自己的判断,看看这是否真的是你最佳的早晨饮品。
安眠药(Sleeping pills)
迈克尔·杰克逊是个天才,他痴迷地关心自己的健康,想成为不朽或至少活过 150 岁,他甚至助长了他睡在氧气帐篷里对抗衰老的传闻。然而他犯了睡眠卫生的根本错误:用药物控制睡眠,这就是为什么他在 50 岁去世。他负担得起最好的医疗建议,然而这位流行天才死于无知。
地球上数百万人服用苯二氮卓类让自己入睡,另一些人把自己喝睡,还有人吸一口大麻。不可避免地,结果都一样:不令人神清气爽的睡眠和白天疲倦。Kripke 博士在其研究中表明,服用安眠药的人更早死亡。那么为什么这么多人犯下用药睡眠的错误?对许多人来说,不令人神清气爽的睡眠总比不睡好。更重要的是,作为社会,我们失去了对一夜令人神清气爽的睡眠能为大脑和身体做什么的真实感觉!所有这些损害都发生在我们本可以用很少的努力就让大多数健康人获得好睡眠的时候:
- 在正确的时间睡眠,以及
- 睡到自然醒来(更多见:Formula for good sleep)。
只要可能,避免诱发睡眠的药物。即便是看似天然的产品褪黑素,也有其缺点。相反,请阅读关于自由睡眠的内容。
褪黑素(Melatonin)
褪黑素是唯一被证明有效的天然安眠药,副作用少。难怪它越来越受欢迎。此外,其抗氧化特性引发了对褪黑素作为抗癌剂的兴趣。在这种兴趣的浪潮中,总有一波廉价的假药冲击市场,尤其是通过互联网销售。这些可能根本不含褪黑素。
褪黑素是一种天然睡眠激素,在松果体(pineal gland)中由5-羟色胺(serotonin)通过血清素 N-乙酰转移酶催化的乙酰化合成乙酰血清素(acetylserotonin),后者随后在 SAM 参与下甲基化为褪黑素。褪黑素在夜行动物和昼行动物中都在与黑暗期对应的昼夜节律周期部分释放。像我们这样的昼行动物,是在光照期活动的。然而,褪黑素只是复杂入睡过程中的中间体。它可以加速入睡,并可以略微提前睡眠相位,然而,它不能按需产生睡眠,它触发的睡眠在结构上常常不同于正常的健康睡眠。褪黑素对睡眠结构的影响,可能是许多使用褪黑素作为助眠剂的人报告早晨不那么神清气爽的原因。对其局限性的解释可能在于,褪黑素在 SCN 下游产生,因此不能被视为影响入睡机制根源的通用睡眠激素。褪黑素沿其独特的 PRC 产生相位移动:
然而,褪黑素诱导的相位移动在多大程度上是褪黑素对 SCN(大部分褪黑素受体所在处)直接影响的结果,又在多大程度上是较早醒来(傍晚给药)的唤醒的相位移动力影响的结果,尚不清楚。无论答案如何,用褪黑素诱发的睡眠不太可能在生理上等同于自然睡眠,因为它绕过了某些昼夜节律控制阶段。我猜测它可以比作在略早的相位睡眠,伴随相应的睡眠结构变化。
高达 0.3 毫克的剂量可将血清褪黑素提升到其天然夜间水平。褪黑素半衰期约 40 分钟,这对定时给药以诱导昼夜节律相位移很重要。高剂量褪黑素(1 毫克以上)的副作用包括认知损伤、困倦、恶心、头痛以及困扰的梦境意象。褪黑素对认知的负面影响是由其对睡眠结构的影响还是对大脑和/或其他组织的直接影响造成,尚不清楚。高剂量可能适得其反,因为它们可能因对 PRC 延迟侧的延长作用(该侧在主观夜晚很早就开始)而产生相位延迟。毋庸多说,出于同样原因,在不眠之夜额外给药会与及时傍晚使用的期望效果相反。
褪黑素可用于纠正 DSPS 中的相位移,然而它不能被视为治愈。要使其效果持续,需要持续给药。停药可能因各种下调问题和内源性释放抑制而实际加重症状。总睡眠时间不增加,而主观警觉性可能实际下降(Sack 等,200739)。褪黑素也被考虑用于 ASPS 的早晨给药,然而考虑到其对认知的影响,这一应用几乎肯定会高度有争议。
对于创造型个人,褪黑素应只在绝对必要时使用(例如为了产生维持社会所需作息所需的相位移)。时机和剂量对治疗效果至关重要,必须咨询合格的专业人士!
尼古丁(Nicotine)
吸烟破坏睡眠、破坏健康,并过早地杀死许多好人。你觉得奥巴马是个冷静理性的人吗?注意他正牢牢地被尼古丁成瘾掌控。尽管他声称恰好及时戒烟以成为总统,传闻说他仍不时偷偷抽一口。
为了好睡眠和好健康,现在就戒烟!
你听说过一种叫”增量戒断(incremental withdrawal)“的戒烟好方法吗?可能没有,这个术语是我现场创造的。然而,我看到许多人用这个方法成功。它是这样进行的:
- 精确计算你每天吸多少。让我们称那个数字为基准计数(Baseline Count)。
- 取 Baseline Count 的 95%,称之为你的当前计数(Current Count)。
- 每天早晨,确保把 Current Count 放进烟盒。按你希望的方式计划,那盒烟必须是你那天吸的全部!
- 你在 95% 水平感觉更好吗?你觉得你能无限期维持吗?如果你对这些问题有强烈感觉,把 Current Count 减少 1。
- 坚持 Current Count,直到你真的有心情再前进一步。记住,把 Current Count 减少超过 1 对你的决心有风险。记住,在减少 Current Count 之前,经历日程中的整组日子(如工作日 vs. 周末)是有意义的。在确定之前不要跳跃。
- 回到第 3 步并留在这个循环,直到 Current Count 降到零。记住,成功的关键是始终对过程感觉良好。你需要感觉进展并享受它,而不遭受太多戒断后果。
我喜欢增量戒断方法,因为它增加了成功的机会。此外,突然完全戒断并非没有风险。戒烟对心血管系统总是好的,然而戒断可能对该系统造成巨大压力,它实际上可能杀死你!我偏爱较少玩弄人体生理危险方面的方法。如果你真的想证明自己的力量,快速通过增量过程而非瞬间戒断。突然完全戒断不仅有风险,还增加复发的机会,这源于不朽的格言:来得容易去得快。更多提示:How to quit smoking?
如果你仍离不开尼古丁,Nicorette 口香糖可能是最简单的非处方方式,以无香烟致癌效应的方式应对成瘾。奥巴马就靠 Nicorette。然而 Nicorette 本身可能比香烟更成瘾,尼古丁的短半衰期可能导致扰乱睡眠的夜间渴望!
记住,吸烟者通常经历浅而不令人神清气爽的睡眠。吸烟者比不吸烟者得到更少的 REM!尽管尼古丁让你感觉更有创造力,没有 REM 睡眠,你的创造力和问题解决能力不可避免地下降。不要被尼古丁注射的短暂效应愚弄!从长远看你会更不聪明!尼古丁通过作用于唤醒区域(包括一个重要的睡眠中心 基底前脑(basal forebrain))的胆碱能受体来改善你的警觉性(见:Why do we fall asleep?)。然而,它也会导致夜间戒断效应,常导致过早醒来。你经常只睡 2-4 小时就醒来吗?如果你戒烟,你可能把这些过早醒来抛在身后。
有趣的是,SuperMemo 用户中只有 4% 是吸烟者(来源)。此外,吸烟的用户花在 SuperMemo 学习上的时间少得多(平均每天约 10 分钟,而通常平均约 30 分钟)。这更多与吸烟者大脑的激素平衡有关,而非吸烟本身。吸烟者只是对 SuperMemo 没有耐心,不太可能是深入的学习者。然而,有强烈迹象表明,戒烟者在学习的毅力上有所改善!这是戒烟的又一个理由!
运动(Exercise)
运动对睡眠有益(Exercise is good for sleep)
如果运用得当,运动是睡眠的恩赐。运动有益健康,而凡是有益健康的也有益睡眠。运动已知能增强深睡眠并促进生长激素的夜间释放,后者被发现通过其对蛋白质合成的影响来刺激记忆巩固。运动、深睡眠和 GH 都与神经发生(即大脑生长)相关。运动后的一夜好眠会导致 BDNF 释放增加和神经细胞增殖增加(例如见:Running increases cell proliferation and neurogenesis in the adult mouse dentate gyrus(Van Praag 等,199969)、Sleep deprivation reduces proliferation of cells in the dentate gyrus of the hippocampus in rats(McGinty 等,200370))。换言之,运动同时构建肌肉和大脑!(Gambelunghe 等,200171)。
睡眠、学习和运动是聪明大脑最好的朋友!
什么时候运动最好?(What is the best time to exercise?)
有不同形式的运动:类型、强度和时机都会影响睡眠。可能的排列组合数量巨大,需要另一篇文章来列举。阅读你喜欢的运动形式,并自己实验!我倾向于在昼夜节律曲线的下降斜坡上运动,无论是在午餐或晚餐前,还是在傍晚早些时候。这个选择是出于为创造性工作保留最高警觉性时段的愿望。此外,峰值警觉性时间不一定是峰值体能表现时间,后者来得稍晚。打比方,大脑比心血管尤其是肌肉骨骼系统醒来得更快。午睡前一餐前的运动可以加深中午小睡,只要有足够的”冷却”期。傍晚晚些时候的运动可能对运动本身最好。耐力和力量在那个时间倾向于达到峰值,而受伤的机会最低(一种理论说这是由于最不分解代谢的睾酮/皮质醇比值)。傍晚早些时候的运动,如果不强度过大或导致受伤,会对夜间睡眠质量产生奇效。此外,傍晚运动似乎是大脑已经结束脑力努力能力时段的良好填充。运动可能比学习更能增加对睡眠的需求。然而,高肾上腺素、竞争性、情绪化或导致受伤的运动如果离主观夜间睡眠太近,可能具有破坏性并降低睡眠质量。此外,有一些例外,它可能不如在更合适的昼夜节律相位运动健康。晚运动可能增加受伤风险,也可能导致心血管系统(本应夜间放慢)的矛盾信号。睡前运动比饭后运动稍不矛盾。这样,傍晚晚些时候的马拉松应被劝阻,而一些体操或适度的健美可能被鼓励。同样,你自己的实验是必不可少的。如果你的运动方式感觉很好,创造性工作不受影响,睡眠很好,很可能你一切都做对了,出于心理原因你更可能坚持运动作息。清晨运动对与睡眠相位问题斗争的人很好。如果你发现难以足够早入睡并因睡过头而需要诉诸闹钟,清晨在明亮阳光下慢跑可以帮助你移动睡眠相位。运动和光是强大的授时因子。自然,这种做法也会导致你在一天中更早耗尽精神能量,这有时正是你需要的,以纠正睡眠相位问题。共和党总统候选人 Ron Paul 承认,对他来说最佳的运动时间是一大清早,他规律地在一天开始时长途步行。这对他有效,因为步行不导致受伤,确实可以很早进行。此外,对政治家来说,这种运动不需要与他的创造性作息争吵。步行很好,例如,可以重申晚上辩论的主要谈话要点。Paul 的例子表明,没有一刀切的规则适合所有人。例如,傍晚晚些时候的运动倾向于延迟睡眠相位,可能对患有 ASPS 的人有帮助。
最佳运动的公式(Formula for best exercise)
如果你以热情甚至亢奋期待你的运动,你就知道你选择了正确的类型、时机和强度。如果你感觉愉快地疲惫,快速入睡,睡眠深,你就知道你做对了。如果你无法拖自己去慢跑,试试步行或游泳。如果水太冷,也许试试室内游泳池。如果游泳不够有挑战,试试团队运动的肾上腺素。或者也许健身房的社会环境更适合你。如果运动加重你的失眠,试试更早,改变强度或运动类型(例如改为不那么导致受伤的)。阅读相关运动提示(补水、防止受伤等)。运动可以也应该令人上瘾。许多人讨厌运动,因为他们从未正确尝试过。如果你是其中之一,再试一次,也许和私人教练一起(一段时间)。人们只有在不知道如何运动时才讨厌运动!
倾听你的身体!运动应让你之前热情,之后知足。如果是这样,你几乎肯定在正确的轨道上。
睡眠对运动有益(Sleep is good for exercise)
至少在健美者中众所周知,睡眠对肌肉生长和修复是必要的。如果你睡眠不足,你的健美努力将被毁掉。肌肉需要睡眠,但我们不因为肌肉的需要而睡眠。为了让机体应对肌肉再生,没有必要关闭中枢神经系统并让一个人一生中三分之一的时间无意识。如果 REM 瘫痪要扮演这个角色,它可以在延髓(medulla oblongata)层面强制执行,而不让我们无意识。如果生长激素分泌要扮演角色,它也可以在与中枢神经系统状态抽象的情况下被上调。睡眠对肌肉再生还有许多其他好处,但没有任何一个单独需要无意识状态。对演化来说,用睡眠进行肌肉再生就像为了修公路而关闭政府一样不合理。睡眠进化以促进休息和再生的普遍信念,来自一旦你睡眠不足就感觉”散架”和”不神清气爽”的感觉。然而,通宵后你感觉崩溃不是由于缺乏睡眠造成的损害,你的状态只是你身体对抗睡眠不足的自身防御。你可以在一定程度上欺骗这些防御,一夜好眠后你的身体似乎就恢复状态了。睡眠剥夺造成的唯一真正损害是对记忆肌理的,不幸的是这种损害不可察觉,普遍的感知是睡眠廉价、可以轻易省去。无论你为记忆还是为肌肉而睡眠,睡眠都是好的。
性爱与运动不同(Sex and exercise differ)
有人注意到我自相矛盾:推荐睡前性爱,又说睡前直接运动不推荐。只需观察到激素上性爱与运动天差地别,性爱中的压力程度应可忽略(至少在稳定和谐的关系中),受伤机会不高等。这种睡前运动我全心全意推荐。
电视(TV)
许多睡眠专家说,卧室只应用于睡眠和性爱,他们也暗示睡觉的地方不应有电视。然而,这一建议似乎源于与睡眠相位问题常常导致的入睡性失眠的徒劳战斗。如果你很早入睡,不够困,那么电视或收音机提供分散注意甚至过早唤醒你(例如恼人的响亮广告)是相当自然的。然而,如果你在正确相位睡眠(例如自由睡眠中),电视声音的效果可能相当不同。在那种情况下,你在真正准备好的时候入睡。如果你播放预录材料,并把定时器设为 5-15 分钟后关闭电视,你可能在为你的睡眠做好事。一夜好眠的重要事项是把一天的所有问题抛在脑后,即便是与你生活相关的愉快想法也可能让你保持清醒和兴奋。同时,来自异国的重复新闻或中等有趣的科学节目可以在 2-4 分钟内有效地哄你入睡,这应是你的健康目标。对成年人来说,一些电视或收音机新闻可以比作给新生儿的舒缓音乐或妈妈睡前给孩子讲的寓言,这甚至已成为书目疗法(bibliotherapy)的一部分。许多人读到睡着,这是个好主意(只要充满激情的阅读不持续到早晨)。有声书可能以不同方式也有效。这是非常个人化的问题,你需要自己测试并避免教条。如果你需要早起并有入睡问题,你可以遵循传统建议。然而,如果你负担得起自由睡眠,你应只在真正困倦时入睡。在那种情况下,要不要电视真的不是问题。你可以获得那 2-3 分钟的新闻,或就在绝对安静中入睡,由你决定。
大麻(Cannabis)
大麻是一种著名的睡眠”疗法”,在 DSPS 患者中尤其受欢迎,他们声称它帮助他们更早入睡。不幸的是,研究似乎表明,大麻改变睡眠结构(例如降低 REM 睡眠比例(Feinberg 等,197572;Fujimori 和 Himwich,197373)),这也可以在戒断时特别清晰的梦境中表达。这导致较低的睡眠效率和给药后可能的过早醒来。由于建议的对褪黑素释放、催眠效应和可能过早醒来的影响,大麻可能看起来像可以稳定 DSPS 中昼夜节律周期的疗法。然而,这种稳定将以睡眠质量和生产力为代价。主观困倦报告似乎表明,大麻吸烟者确实醒来时远不那么神清气爽。此外,他们在戒断时经历异常高的能量水平和丰富梦境,这表明该药物对睡眠质量的负面影响。
撇开吸食大麻的其他潜在负面健康影响不谈,它绝对应在”保护区”(即睡眠前数小时)避免。在这方面,它与酒精或苯二氮卓类没什么不同,后者也众所周知会影响睡眠结构和睡眠效率。在所有这些情况下都适用同样的规则:对待你睡前的大脑,不比你对待创造性工作前的大脑更糟。凡是对创造力有害的,都可能对睡眠质量有害。
Buzsaki 博士在一个采访中说:“时机和网络同步是所有皮层计算的本质,皮层网络的时机能力反映在它们产生的节律中。我们已经表明,海马体组件同步的退化(例如由大麻活性成分诱导的),定量地反映在场节律中。反过来,海马体振荡受损的程度与记忆表现的退化相关。[……]振荡构成一个稳健的表型,可靠地’指纹识别’个体,并预计在大多数精神疾病中改变。通常,这种变化在睡眠中最明显。“
性爱(Sex)
性爱对睡眠有益,然而把性爱当作”安眠药”可能对性爱本身不太好。由于昼夜节律原因,早晨性爱应是最佳的(在自由睡眠条件下)。睾酮在早晨达到峰值。然而,性爱是强大的催眠剂,早晨性爱可能削弱早晨警觉性。另一方面,睡前性爱可能有助于你更快入睡。傍晚性爱可能不那么”高效”,你更疲倦,也许没心情,但性爱是极好的催眠剂!性爱是非常个人化的事,但我相信,创造型人在睡前或午睡时间”性爱运动”后表现更好。
如果你进行不以生育为目的的性爱,性爱对睡眠的正面影响可能是你忠实履行夫妻义务的头号借口。这里也推荐坚持单一伴侣。长寿研究表明,健康稳定的单配偶性生活是强大的预期寿命决定因素之一(即便在这种情况下,单配偶应与滥交而非多偶相对立)。虽然单配偶性爱总体对睡眠有益,但与你新恋人的性爱可能实际上扰乱睡眠。除了健康剂量的内啡肽,它还会提高你的儿茶酚胺,可能碎片化睡眠周期。出于同样原因,滥交性爱也可能无法扮演预期的催眠角色。
许多高度创造性的人选择与伴侣分床睡。这种方法可能削弱家庭凝聚力和性生活,然而相当可以理解,尤其是那些喜欢两头点燃蜡烛的人。共同睡眠在健康上可能是更好的选择,只要它不影响睡眠质量。此外,双方应在相似时段睡眠并放弃闹钟。这是一个艰难的个人选择,需要在健康传统与追求生产力之间取得平衡。我不认为我能推荐一种选择而非另一种,它太复杂也太个人化了。
饮食(Diet)
我们吃的食物影响我们的警觉性和睡眠倾向。然而,食物的作用被大大高估了。例如,用基本食物显著影响昼夜节律周期非常困难。更重要的是用餐时机。内稳态困倦可以通过一些食物增强,尤其是大量摄入时。然而,这高度个体化。例如,你的葡萄糖耐受性将决定富含葡萄糖食物的影响。你代谢酒精或咖啡因的能力将决定这两种常摄入的精神活性物质的影响程度。你的食物不耐受和食物过敏也可能有大影响。你当前的饱腹状态、补水、热量需求等也起作用。“睡前一杯温牛奶”等古老建议在帮助你睡好方面只起边际作用。需要单独一篇文章来描述所有细微差别和可能的相互作用与协同。因此,我将本节限于以下基本经验法则助记:
- 凡是对健康有益的,也可能对你的睡眠有益(至少长期)。健康的身体确保健康的睡眠。咨询你最喜欢的健康饮食书籍或网站,持续学习!你几乎需要背下一本食物百科全书才能真正理解不同饮食选择的所有细微差别。
- 如果你偏胖,避免在清醒日的后三分之一进餐。出于激素原因,这将帮助你控制体重,并帮助你睡得更好。
- 即便你在减肥计划中,创造性工作前你需要能逐渐向血流释放葡萄糖的食物,以便你在高挡位工作时使用。
- 避免在清醒日的后三分之一摄入酒精和咖啡因。
你为好睡眠的最佳饮食,大致就是对你健康和长寿有益的饮食。
健康饮食的变化不太可能对你的睡眠有重大影响。你可能只想留意咖啡因、酒精、外来草药产品、毒素和所有对神经系统有实质性影响的物质。否则,饮食应只对睡眠需求、昼夜节律模式、内稳态困倦(咖啡因是主要例外)、睡眠阶段进展(酒精是主要例外)或睡眠的神经效率有轻微影响。原因与许多其他稳态情况相同:机体努力在所有系统中保持稳态平衡。稀有食物、草药制剂、药物干预等总可以改变或不平衡内部平衡,但标准的健康饮食不太可能这样做。阻止人心脏需要非凡的营养错误,停止基于基因的生物钟更难。
素食(Vegetarian diet)
关于纯素或素食饮食与睡眠的关系有很多说法。草食动物睡得更少不是真的,因为有许多例外(有些草食动物的睡眠是短睡眠肉食动物的三倍)。然而存在一种相关性,即随体重增加,草食动物睡眠时间的减少比肉食动物更快。换言之,像长颈鹿这样重的草食动物确实睡得很少。这种相关性可以用代谢变化解释,但也可能反映不同的生活方式。捕食者可能吃一次然后花很多小时消化,大象整天不停咀嚼以维持能量需求,而瞪羚需要最大化警觉以抵御长时间睡眠的狮子的攻击。饮食类型(草食 vs. 肉食)与睡眠长度之间的相关性,把睡眠习惯与物种的饮食习惯联系起来,而非个体的饮食习惯。虽然人类是杂食的,你不会通过强制新的饮食习惯变成草食动物,并据称成为短睡眠者。
睡眠与葡萄糖代谢(Sleep and glucose metabolism)
虽然健康饮食的变化对睡眠质量影响不大,但睡眠对代谢有强大影响。睡眠剥夺研究告诉我们,充足睡眠对健康的葡萄糖代谢尤为重要。睡眠剥夺、轮班工作和时差都促进肥胖和2 型糖尿病的发展。可能的原因是,睡眠剥夺在相同热量摄入下降低瘦素并增加饥饿素(Knutson 等,200774)。这两种激素控制食欲并影响体脂水平的稳态设定点。在睡眠剥夺中,我们倾向于吃得更多,并在会增加体脂百分比的点上达到饱腹。以热量计,这些变化可能相当剧烈。把睡眠减半可能使食物需求每天增加 1000 千卡,还有迹象表明食欲转向大幅偏好高碳水食物,这只会放大问题。最后,睡眠剥夺让你变懒,你更不可能消耗那些额外热量。
获得充足睡眠帮助你保持苗条!
更多内容见:The Dream Diet: Losing Weight While You Sleep。
禁食(Fasting)
大多数营养学家会告诉你,如果避免在一天后三分之一吃大餐,减肥更容易。另一些人声称这是迷思。流行病学研究一如既往地不提供明确的证实。斋月禁食(Ramadan)似乎有利于减肥,尽管夜间进食。对照研究也根据设计提供看似矛盾的结果。傍晚禁食可能有益的信念可能源于 1970 年代,当时减肥计划被证明在食物在一天前半部分(而非后半部分)食用时更有效(讨论见:Weight Loss is Greater with Consumption of Large Morning Meals(Keim 等,199775))。
我相信,如果你亲自尝试傍晚禁食,你会很快发现它可以对你的睡眠、其恢复力、你的减肥目标、你的早晨能量等产生奇效。除非你是那种没有夜间去冰箱或至少傍晚零食就无法睡眠的贪婪型,你还会注意到,由于昼夜节律和心理原因,一旦你掌握了诀窍,傍晚禁食相当容易维持。禁食促进饥饿素的释放(Bloom 等,200076),这有助于夜间生长激素释放的总体增加(Norrelund,200577)。大多数生长激素释放发生在夜间早期的较深 NREM 睡眠阶段。这种夜间释放部分负责早期睡眠的合成代谢模式,帮助你避免腹部肥胖、强化骨骼、重建肌肉、肌腱、韧带和其他日间磨损组织。激素上,傍晚禁食产生的效果类似于总体热量限制,后者已被证明可延长小鼠寿命。寻求青春过往的老年人可能诉诸生长激素注射。傍晚禁食结合健康的自由睡眠,绝对是更健康、更简单的选择。亲自试试,如果有任何疑问,请写信给我。
如果你是失眠症患者或患有 DSPS,你也应考虑傍晚禁食作为可能帮助维持健康睡眠作息的因素。见:Curing DSPS and insomnia。另一方面,如果你受早醒和短夜困扰,你可能违背传统营养师建议,听听 Seth Roberts 的相反说法。Roberts 发现跳过早餐帮助他维持健康的睡眠相位(Roberts,200478)。数千人遵循 Roberts 的建议,却没有意识到他们中大多数可能处于相位障碍谱系的 DSPS 端,而他的建议虽然研究充分、非常适合他,但在他们自己的情况下可能产生相反效果。因此记住,你的禁食选择以及其他影响睡眠的生活方式改变,必须选择以适合你的睡眠时型。
用饮食对抗时差(Combating jetlag with diet)
有一些证据表明,大鼠可以借助 DMH 把周期驯化到食物,然而,用饥饿对抗时差只是一个理论概念。SCN 节律不能在轻微相位移之外被塑造,SCN 和 DMH 之间失去同步(如果人类可能的话)不太可能对健康有益,尤其是人类似乎没有进化出让睡眠服从食物可得性时机的机制。如果你碰巧在用餐时机对抗时差方面有任何成功,请告诉我。
学习(Learning)
睡前很少有像一剂繁重学习那样令人疲倦的事。然而,一位领先的睡眠专家 Dement 博士在他更好的睡眠指南中建议:“睡前避免繁重学习或电脑游戏,它们可能令人兴奋。” 这一建议需要轻微修正。毫无疑问,电脑游戏令人兴奋,应避免。然而,“繁重学习”可能有许多形式。如果你为考试学习,这带来考试本身的压力形象,确实可能令人兴奋。如果你学习一个迷人的学科,垄断你的思维,它也可能令人兴奋。同样,在明亮照明的房间里学习可能减慢入睡下降。然而,如果你提取纯粹的学习过程,剥离压力联想、光线、社交等方面,你会得出不同的处方。
学习应帮助你入睡(Learning should help you sleep)
你在给定的一天学得越多,学习更多的能力就越低(见:Learning overload)。因此,你学得越多,越快会严重困倦。然而,在昼夜节律主观夜晚到来之前,你无法睡好。这意味着你只能略微提前入睡时间,例如 20-60 分钟。你无法用学习产生多小时的相位移!
学习与困倦的内稳态组分相关,可以促进睡眠。
如果你想用学习作为为睡眠疲倦的方式,并且你不介意学习过程效率较低,以下是建议:
- 选择一些不令人兴奋的学习材料(例如你的法语词汇可以通过测试,除非你计划一次激动人心的巴黎之旅)。
- 减去高优先级材料(在困倦状态下学习可能对该子集的学习过程产生负面影响)。
- 确保你的显示器和房间不太亮,否则你可能影响睡眠相位,让第二天的睡眠更难。
记住,在困倦状态下学习实际上是违反学习卫生的。科学尚未结论性地回答这对你的长期记忆是好是坏的问题(Wozniak,200279)。
学习会增加总睡眠吗?(Does learning increase total sleep?)
尽管你常听我说学习增加睡眠需求,我一直无法用 SuperMemo 数据证明这一事实!我只是重复其他科学家一直说的话。学习确实应该增加睡眠中神经优化的需求,然而,这也可以通过提高处理强度(例如提高 REM 睡眠密度)来完成。繁重学习不一定增加睡眠长度。学习也可能像运动,对基线需求贡献不大。如果你不用教科书学习,你仍然通过注意事物、思考、与人交谈来学习。如果你不运动,你仍然燃烧大量卡路里。似乎更容易证明繁重运动导致更长睡眠,而非证明繁重学习增加总睡眠。我已经能够表明学习对内稳态困倦有贡献,因此它应该有助于更早入睡和更长睡眠。然而,我仍无数据表明这一点。更多内容见:How learning affects sleep?
睡眠与学习(Sleep and learning)
睡眠长度(Sleep length)
最佳睡眠长度(Optimum length of sleep)
在这个繁忙的现代世界,每一分钟时间都显得珍贵。对一些人来说,瓶颈资源是时间(不是金钱、物质资源、人等)。时间成为限制因素,每个人都在寻找疏通生活的方法。由于睡眠占我们生命的三分之一,一个广泛的无知解决方案是削减睡眠以节省更多工作时间。这在短期内可能对需要腿或手臂多于大脑的人有效。对那些以大脑为主要工具的人,这肯定会适得其反。即便对那些相信自己能在有限睡眠下完成更多的人来说,生活也一定像一场没有快乐阳光的赛跑。从全球价值看,一个神清气爽的大脑产生的单一创造性洞见,可以等于数千人时的繁重劳动。数百万在第一次世界大战前线浪费的年轻生命,其有意义的贡献可能还不如 Tim Berners-Lee 几小时的编程。想象所有那些生命用于更有生产力的追求!通过削减睡眠,你削弱了自己做出有意义创造性贡献的机会(除非自然地,你的有价值使命没有一些睡眠牺牲就无法完成)。
推荐的 8 小时睡眠(Recommended 8 hours of sleep)
睡眠研究者常寻找推荐的睡眠量。使用调查或实验室睡眠数据,他们常得出每晚 8 小时睡眠的推荐。然而,这一推荐打开了问题的雷区。为了获得 8 小时,一些人可能希望过早入睡,从而加剧失眠和相关压力。每晚 8 小时的推荐也被一些研究者嘲笑,他们传播睡眠像食物、即使不需要我们也总想要更多的错误主张。
睡眠的节能理论公然错误。睡眠的好处不同于食物的好处,不能提前积累,获得超过必要的睡眠没有演化优势。一个月过度睡眠后的通宵,和仅仅 2-3 晚好睡后的通宵一样痛苦。节能微乎其微,大脑在某些睡眠阶段实际可能比处理复杂任务时消耗更多氧气。尽管狮子在缺水缺食时可能每天睡 20 小时,人类在正常情况下,只能在睡眠剥夺期、或在错误昼夜节律相位睡眠、或经历健康问题时暴睡。理论上,神经网络优化可以从一些额外睡眠中受益,然而大脑似乎并不渴望那种额外优化。它的控制机制设定让睡眠每天持续有限时间,即便我们努力获得更多睡眠。不仅没有好处,睡得太多还有巨大成本:我们在睡眠中最脆弱、最无防御。尽管睡眠可以压缩(证明它不完美高效),但没有自然健康的睡眠压缩方法。最好的睡眠是在所有昼夜节律、内稳态、基因和神经机制在正确时间按既定课程同步运行时实现的。这只能在自由睡眠中实现。凭借数十份 SleepChart 提交,我可以轻松证明,一旦坚持规律睡眠作息,自由睡眠的总量下降并变得相当稳定。在示例图中,一位 DSPS 受试者自由运行睡眠,平均每晚获得同样规律推荐的 7.9 小时令人神清气爽的睡眠,尽管在自由运行之前,她确信自己需要 9 小时,且即便 9 小时她整天仍会疲倦:
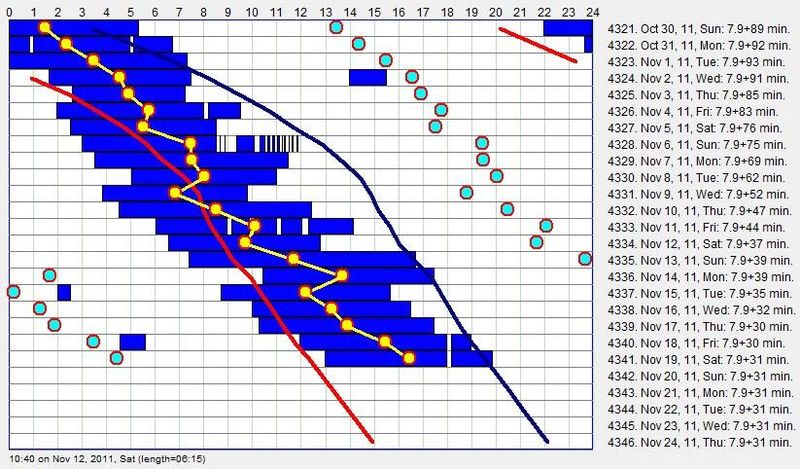
用多相睡眠节省时间(Saving time with polyphasic sleep)
Uberman 睡眠作息是为了获得更多一天中的小时而提出的。多相作息在理论上非常有吸引力,许多人尝试后在一周或一个月内放弃(取决于承受精神痛苦的能力)。那些试图适应任何不自然作息的人,将承受无法言说的精神折磨。多相睡眠者用闹钟调节睡眠,直到达到崩溃点。人类自我实验的豚鼠在崩溃点附近崩溃成 5-8 小时的生命拯救性睡眠,然后带着罪恶感恢复多相作息。那种罪恶感用”周末休息”、“再充电”、“额外睡眠”等开脱性术语来平息。多相实验者可能睡得更少,但他们的智力表现将被大幅削弱。一些多相睡眠理论基于身体可以适应任何睡眠节律的错误前提。研究者试图找到一个自然的多相节律,以最小化少睡的痛苦。为此,他们研究了昼夜节律的相位响应曲线,其中各种影响睡眠的因素的影响被显示为向前或向后移动睡眠作息。显而易见的结论是,我们可以相当无痛地把主要昼夜节律困倦时间一点点移到期望方向。然而,健康的正常个体无法把节律切成期望数量的片。单相或双相睡眠是健康个体的常态。双相睡眠很少由两次主要睡眠事件组成,通常采取主事件(夜间睡眠)加次事件(午睡)的形式。伟大的打盹者在感觉需要时打盹,常常能准确预测何时以及需要多少。如果你想最小化睡眠时间并最大化学习结果:自由运行睡眠。多相睡眠不是答案。扔掉闹钟!
最小化睡眠时间(Minimizing sleep time)
所有用闹钟控制睡眠的形式都会增加对睡眠的总体需求。这意味着:
如果你使用闹钟,要么:
- 你会睡得更长,或
- 你会感觉更悲惨。
人为睡眠作息会大幅降低你的脑力能力。健康个体在正常条件下会发现,主要睡眠事件后 4 小时难以入睡,除非该事件被闹钟不自然地切断导致睡眠剥夺。
最短的健康睡眠是在自由睡眠中实现的!
在自由睡眠中,一旦你知道自己的平均睡眠时间和最佳醒来时间,就一丝不苟地坚持它。如果你的睡眠不规律,用 SleepChart 找到你的最佳值。这样计划你的一天:如果困倦来得更早,你可以眨眼间上床;如果困倦没按时到来,你可以忙于一些低优先级、有助于睡眠的活动,让你疲倦直到正确的睡眠时间。
睡眠控制系统中的低效(Inefficiencies in the sleep control system)
许多因素可能增加睡眠需求(如学习、运动等),或把睡眠控制平衡移向有利于睡眠而非清醒(如脑损伤、感染、中毒、体温过低等)。外部因素的影响可以用来说明睡眠控制系统的一定程度低效。由于夏天比冬天白天长,我们在冬天确实倾向多睡一点。冬天似乎没有增加对睡眠神经功能的需求。如果我们冬天睡更多而没有更多睡眠的生物需求,那么似乎我们要么冬天睡得比需要的多,要么夏天睡得比需要的少。由于睡眠主要由昼夜节律和内稳态睡眠倾向控制,而昼夜节律组分强烈受光影响,光照水平的变化会导致睡眠时长的变化。那么可以想象,我们冬天睡得更低效(以单位时间的神经效应计)。同样,夏天睡眠可能不那么具恢复性。脱离文明影响的因纽特人冬天每天多睡几小时。Jim Horne 博士说,在某些情况下我们可能睡得比真正需要的多,这是对的。然而,当他把睡眠比作进食,使一些人相信睡眠限制可能有益(类比热量限制)时,他走得太远。总之,我们需要认识到睡眠控制机制不完美,然而我们还没有任何改进生物演化所给予的、人工且认证的方法。自然自由睡眠仍是实现健康、令人神清气爽且最短睡眠的最佳方式。
SuperMemo 用户的睡眠长度(Length of sleep among users of SuperMemo)
一项对 SuperMemo 用户的调查(SuperMemo World,199480)显示,平均学习速度为 243 项/年/分钟。那些睡眠少于 7.5 小时的用户以 240 项/年/分钟的速度学习,睡眠超过 7.5 小时的用户以 256 项/年/分钟学习。睡眠量、吸烟和运动与学习速度的相关性较弱。28 岁或以下的学生以 264 项/年/分钟的速度学习,而 28 岁以上的以 179 项/年/分钟学习。然而记住,在另一项研究中已经表明,好学生学习更慢(!)(Gorzelanczyk 等,199881),因为他们在给分时更自我批评。
睡得少的人活得更长?(People who sleep less live longer?)
2002 年 2 月,新闻头条惊呼”睡得少的人活得更长”。这场骚动的原因是加州大学圣地亚哥分校研究人员的一项大规模研究,他们发现,平均每晚睡眠 8 小时或以上的人在 6 年研究期内死亡的可能性,比睡 7 小时的人高 15%。这项研究为我们对睡眠习惯的知识做出了宝贵贡献,但大众媒体放大的结论不仅错误,而且危险!如果你决定今天削减睡眠以活得更长,你肯定会达到与期望相反的效果。是你的身体(实际上是大脑)最清楚你需要多少睡眠,这可能是 5 小时也可能是 9 小时,我们在这一方面差异很大。用闹钟削减睡眠没有值得注意的好处,而危险有充分记录。媒体从 Daniel Kripke 博士团队研究得出的错误结论,来自典型的因果混淆。不是长睡眠有害,更多是健康状况不佳可能增加睡眠需求。推向极端,昏迷和卧床患者会偏向此类统计。睡眠质量差、有呼吸暂停的人更可能赖床并报告长夜。谱系的另一端是拥有健康、良好睡眠习惯的人,常常仅 5 小时就感觉神清气爽,并在闹钟时间前自然醒来。除了对健康有不良影响,睡眠剥夺是交通事故的主要原因,并对国家的创造潜力造成不可估量的损害。即便设计糟糕的警觉性测试也不太可能证明你的睡眠质量,这是因为压力激素常常掩盖睡眠剥夺。然而,如果你在人为缩短睡眠后尝试用 SuperMemo 学习,你会看到你的回忆变差,压力激素可能改善你的警觉感,但它们会以专注和记忆为代价。在消除压力、兴奋剂、抗抑郁药、安眠药等条件下,获得身体所需的所有睡眠,你将达到最佳健康。大众媒体放大器很可能产生混淆和负面涟漪,未来数年继续回响!
一如既往,一些研究似乎上了头条,而更彻底的荟萃分析却没有。在这种情况下,可能是自我安慰的想法”即便我在睡眠剥夺中感觉悲惨,失去睡眠实际上可能延长我的寿命!“如果我们审视关于睡眠长度与长寿之间联系的文献,我们可能会注意到,非常短的睡眠和非常长的睡眠都与更短的寿命相关(Cappuccio,201082、201083)。Francesco Cappuccio 教授说得好:“虽然短睡眠可能代表不健康的原因,但长睡眠被认为更多是不健康的指标。” 如果你取人口所有成员的最佳睡眠量的平均值,你会得出一个对特定个体无意义的数字。在睡得比平均更长或更短的子群体中,长寿可能降低。然而,对于每个个体,最佳数字是身体需求建议的数字。如果是 4 小时或 10 小时,只要数字来自不受人为控制的自然睡眠,就关系不大。此外,那个数字会日复一日地变化,在激动人心的约会前会更少,在繁重运动的一天后会更多。没有人应该担心每天睡 4 小时或 10 小时,只要他或她自然入睡、自然醒来、感觉神清气爽。
最佳睡眠量因人而异、日复一日变化,最好通过不受人为控制(如闹钟、安眠药等)的睡眠来确定。你可以通过尝试自由睡眠来最佳地确定你的睡眠需求。
关注发病率并问”你每晚睡几小时?“的流行病学研究问了一个错误的问题!它们应该问:“你是否人为修改睡眠的时机和长度?” 只有那种问题会告诉每个人,人为控制睡眠增加发病率,而实际睡眠长度基本无关。
调查睡眠长度与长寿之间的联系,并不比我自己的将睡眠长度与学习质量联系起来的失败尝试更有用。在学习的情况下,短睡眠由于睡眠剥夺对注意力、回忆和巩固的影响产生糟糕结果。然而,长睡眠也产生糟糕的学习,因为它通常是健康或睡眠控制系统出问题的指示(例如在错误相位睡眠,或代偿先前的亏欠)。问我们需要多少睡眠,与问我们需要多少卡路里没什么不同:它取决于我们的体型、当前脂肪水平、卡路里消耗和许多其他因素。
Jim Horne 和 Daniel Kripke(Jim Horne and Daniel Kripke)
Jim Horne 和 Daniel Kripke 是两位睡眠研究者,他们在睡眠像食物、我们可以摄入过多这一突出主张上似乎与该领域其他人对立。他们甚至设想了这样一个概念:如同热量限制,睡眠限制可能延长寿命!我提到了 Kripke 博士常被错误解读为”短睡眠延长寿命”的研究。并非长睡眠者更早死亡,流行病学研究的明显解读是,更病的人常常睡得更长。与热量限制的类比,与提出可能有某些未知好处的”清醒限制”一样站不住脚,尤其是在葡萄糖代谢的激素谱中,清醒限制更类似于热量限制。获得过量睡眠没有演化优势,因为睡眠不能像脂肪那样积累(见:Excessive sleeping)。打个比方,如果我们把睡眠比作垃圾回收,提前收集垃圾没有好处。自由睡眠作息的人很快减少对睡眠的总需求,并比各种形式的调节作息睡得更少,他们的精神能量自然高得多,尽管睡得更少。我相信把长睡眠比作过度进食尤其有害,它向青少年和学生发出错误信号,对他们来说脑力表现决定未来。Horne 和 Kripke 博士主要担心的是,坚持 8 小时睡眠价值的研究者采用的”恐吓战术”可能加重失眠和与”睡不够”相关的压力。这些是合理的担忧,但很容易把平衡摆向另一端,而存在一个黄金中庸:自由睡眠帮助人们获得恰好所需的睡眠量。为了寻求一些平衡,让我挑剔 Horne 和 Kripke 博士的一些陈述和假设,从自由睡眠的角度对睡眠需求投下一些光。
Horne 博士(Dr Horne)
以下是 Horne 博士一些持续减损睡眠价值和力量的陈述:
- “我们需要的睡眠量是白天不困所需的量。” 很难不同意。然而,我会把这一陈述升级到信息时代状态:“我们需要的睡眠量是大脑所要求的。” 这是比”不困”更高的标准。毕竟,在田里干活、在公园散步或打篮球,感到精力充沛不需要太多睡眠。而要达到写作、编程、学习、研究等智力最佳状态,需要更多高质量睡眠。
- “整夜连续睡眠是相当现代的发明。” 这完全错误。许多物种睡眠长达多个不间断小时,这最肯定是早期人类的习惯。自然的睡眠中断,如夜间醒来或分段睡眠,与闹钟造成的强制中断相去甚远。为特定目的醒来可能在史前人类社会就被发明,然而,从来没有频繁使用那个”发明”的迫切需要。几个世纪以来,连蜡烛都只有富人能用。腓特烈大帝童年可能被炮声叫醒,但他的同时代人庆幸地没有疯狂独裁、军事痴迷的父亲。我们的祖先从未对睡眠多加注意,他们就是有很多睡眠,而且从来不是失眠者论坛上现在充斥的讨论或焦虑的主题。闹钟和碎片化睡眠作息以及睡眠问题的真正出现,可以追溯到电照明的早期,它把现代世界与自然的日周期断开。尽管闹钟不新,但它们只在上一世纪大规模使用。
- “关于闹钟有很多危言耸听。你会听到一些有趣的想法——我们应该被光或在我们睡眠模式的某些点’自然’叫醒。” 被光叫醒远比被闹钟叫醒健康。然而,晨光对实现顶级睡眠质量并非必要。见一位八旬老人在无晨光情况下的完美睡眠案例。在最浅睡眠阶段被叫醒远比 15-25 分钟后深睡眠开始时被叫醒健康。大多数人经历过这种差异,可以轻易列举几个表明睡眠破坏对健康负面影响的症状。如果我是关于闹钟的”危言耸听者”,那是因为我足够了解睡眠生理,知道没有闹钟的生活应是基本人权,尤其是在年轻时。
- “失眠实际上不是睡眠障碍,而是清醒侵入睡眠的障碍。” 在其他方面健康个体中,失眠的典型原因是糟糕的睡眠时机。需要早起的人在相对于生物 clock 过早入睡,就是无法入睡。这不是清醒侵入睡眠,而是人希望把睡眠强加于清醒。现代作息和期望是绝大多数情况下问题的根源。
- 在批评用于研究睡眠剥夺的心理测试时,Horne 说*“测试只是测量对无聊的耐受性,它随睡眠剥夺而缩短。拿走刺激,动物就会入睡。”* 创造性工作并不总是提供足够的兴奋来对抗睡眠剥夺。困倦的创造性大脑的敌人不是无聊,而是脑力努力本身。没有任何程度的无聊能让一个休息良好的大脑入睡。 然而,睡眠剥夺中的脑力努力可能等同于折磨。工业化国家的数百万学生每天经历这种折磨。如果我们想测量睡眠剥夺对认知的影响,我们需要既刺激又消除兴奋变量的测试。对无聊的耐受性不随睡眠剥夺缩短!真正厌恶精神虚无的是新鲜的创造性头脑!在睡眠剥夺中真正缩短的是我们在缺乏唤醒刺激时保持清醒的能力。无聊不诱发困倦,大脑中没有已知的”无聊核”会对缺乏刺激做出放电反应(除了通过自然睡眠控制系统的去抑制)。即便那个核被识别,它更可能形成唤醒系统的一部分。儿童被剥夺刺激时可能变得暴躁或攻击性。演化使我们聪明,因为我们寻求行动和低熵状态。无聊揭示了潜在的睡眠剥夺。 任何在无聊讲座上打盹的人,必然只是夜间没睡够。神清气爽的头脑反而会飘向更有趣的想法,如令人兴奋的项目、当天的问题或附近有吸引力的同学。我的印象是,提起无聊对清醒影响的睡眠研究者,必须是从自己睡眠剥夺的状态推断。顶级质量的创造性头脑不需要肾上腺素或多巴胺的鞭策就能专注于创造性追求。我们都应以敏锐清晰的头脑为目标,而非仅仅是”不困”的头脑。
- Horne 博士把因纽特人作为人类过量睡眠能力的例子。因纽特人冬天能睡长达 14 小时。生活方式和环境确实会影响睡眠长度。你可以责怪我们的生物学没有充分利用可用时间,但对此无能为力,所以我们不妨享受完整的夜间睡眠。当不规律的睡眠者声称每晚需要 8-9 小时睡眠时,如果”睡眠浪费”理论正确,人们会预期他们在自由睡眠作息上会达到 10-11 小时。相反,在自由睡眠中他们常常降到每晚 6-7 小时,并声称感觉比以往更好!至于因纽特人,那些额外小时部分可能来自有记录的冬天最小化寒冷暴露的愿望,其中大部分”睡眠”可能是 Wehr 的分段睡眠(Wehr 等,199284)。所以我们需要用多导睡眠图来看那些就寝时间有多少是实际睡眠。另一方面,夏天睡得少可以轻易用以下事实解释:在明亮光线下午夜不易入睡,并且从睡眠另一端有早起打猎、担心可能睡过头的压力。我们知道睡眠控制系统不完美,实际睡眠需求可能被唤醒刺激掩盖。反过来,北极夜晚可能有利于悠闲的睡眠,可能超过机体的实际需求。然而,我们不能肯定地说,是缺乏刺激使因纽特人浪费地睡得更长。我们确实知道,额外的夏季刺激使因纽特人从睡眠中获得的价值低于冬天。要解决这个问题,我们需要在夏天和冬天应用适当的认知测试并比较两者,尤其是比较回忆的昼夜节律峰值。这比回忆平均值更重要,因为我相信创造型人主要在脑力峰值时刻改变世界,即从富含梦境的令人神清气爽的夜间睡眠醒来后不久。在我自己的工作中,我没有发现睡眠长度和学习质量之间的太多相关性。自由睡眠在冬天可能略长,然而,夏天较短的睡眠常被较长的小睡补偿。平均而言,认知表现似乎相当。SleepChart 日志强调反驳了 Horne 博士关于我们喜欢浪费睡眠的主张。自由睡眠减少总睡眠,因为它提高了效率!
- “美国的研究者倾向于不同观点。他们断言,因为我们大多数人可以延长日常睡眠,我们必须需要这样做。这意味着那些工作日似乎满足于 7.5 小时、周末享受 9 小时的人,不知不觉地长期被剥夺,实际上每天需要 9 小时。证据据说来自白天困倦的许多人,以及工作时甚至更糟糕地开车时睡着的众多报告。” Horne 博士对 8-9 小时睡眠推荐的恼怒可以理解,毕竟人们有不同的睡眠需求,推荐可能加剧失眠。然而,周末比工作日睡得更多是睡眠剥夺的明确指示。与工作相关的压力或激情可以轻易掩盖剥夺,那个面具在周末脱落。很难说工作日 7.5 小时加周末 9 小时等于 7.8 小时最佳,还是 8.5 小时最佳,尤其是这些数字即便对非常规律的睡眠者也会日复一日变化。每个人都应在自由睡眠条件下自己确定需求,由于工作周带来的睡眠债,这一条件无法通过周末睡眠来近似。
Kripke 博士(Dr Kripke)
以下是 Kripke 博士一些低估不受打扰睡眠重要性的陈述:
-
“似乎没人确切知道我们应该睡 8 小时的想法从何而来。我猜只是从某人的奶奶传下来的。” “8 小时迷思”并非来自奶奶,而是来自调查,其中许多人声称获得 8 小时睡眠后功能最佳。实际完美数字可能更少,因为常见的知觉错误。工作周睡眠不足的人常在周末补偿,那时他们确实睡得更多以弥补各种睡眠亏欠。那个周末睡眠可能是 8 小时或更多,但这并不意味着那是实际需要的。同样的人在自由睡眠作息下可能惊讶地发现,他们实际只需 6-7 小时。在双相作息上降到每晚 4-5 小时并不罕见,其余睡眠需求由时机恰当的午睡小睡满足。我的小样本估计似乎表明,人们高估睡眠需求 10-15%。Carskadon 博士的研究表明,8 小时对睡眠需求明显增加的青少年可能是良好估计。我自己对 SleepChart 提交的分析也表明,对青少年,8 小时是扎实的推荐!该数字也常出现在其他出版物中(Dinges 等,200085)。当 2011 年头两天的软件故障让 iPhone 闹钟静音时,数千人意识到自己睡过头的能力,不是多睡 10-30 分钟,一些愤怒的博主记录了额外 6 小时睡眠,说明了睡眠剥夺的惊人程度。当奶奶建议 8 小时睡眠时,她们离目标不远!
-
Kripke 在《Sleep》2004 年 2 月的社论中写道:“[流行病学研究的结果] 证伪了广泛流传的最佳睡眠至少 8 小时的假设。” 所有流行病学研究都以这样或那样的方式为我们的知识做出贡献。然而,关注发病率并问”你每晚睡几小时?“的研究是浪费时间!就像关于身高对死亡率影响的研究,我们预先知道它将提供 U 型结果,与我们的生活方式建议无关。正确的问题应是”你是否用闹钟缩短睡眠?“或”你是否在困倦时仍推迟睡眠?“或”你是否在不困时仍试图早睡?“简而言之,这些问题应问:“你是否人为修改睡眠的时机和长度?” 那些研究将明确告诉每个人,人为控制睡眠是致命的,睡眠长度基本无关!更多内容见:People who sleep less live longer?
-
Kripke:“挪威 Hordaland 研究的近期结果显示,报告每晚睡眠少于 6.5 小时的工作者没有遭受残疾的显著增加。那些报告超过 8.5 小时的人,比睡 6.5-7.5 小时的人成为残疾的可能性高出一倍多。这项研究的建议是,如果有任何影响的话,在床上花更多时间可能倾向于增加残疾。” 我们已经知道,平均而言,如果人们睡了一定小时数,或如果他们的身高略高于平均,他们活得最长。但这一陈述似乎暗示反向因果。试图在床上花更少时间对长寿的好处,就像试图把超过某身高的人缩小一样。健康人睡得更少,但那些缩短睡眠的人更不健康!
-
Kripke:“我们知道,每次少于 8 小时的闭眼并不会让我们遭受可怕的智力或记忆丧失。我们刻意选择睡 7 小时或 6 小时,觉得拥有额外时间的实际好处超过了睡眠少于 8 或 9 小时可能带来的任何困倦。” 听到一位睡眠研究者接受用睡眠交换”实际好处”的做法,真让我惊讶!智商的丧失可能不可怕,但道路上的死亡人数绝对是。用睡眠交换”实际好处”有神经和健康成本,一位不可替代的医生或消防员可能做这种交易,但他们应始终记住,他们为他人牺牲了自己的创造力、大脑和健康!
-
在与 Stickgold 博士由《经济学人》组织的辩论中,Kripke 反驳 Stickgold 关于睡眠不足是交通事故原因的主张:“人们声称白天困倦导致道路事故,而事实是与睡眠相关的道路事故率在深夜上升。深夜驾驶造成路上大多数困倦。不寻常地熬夜的人,如夜间被叫值的医生,有额外的驾驶风险和犯错风险,但这并不证明普通人群的规律睡眠模式太短。” 当两位伟大的研究者在可以轻易验证的事实上分歧时,我总可以怀疑专业或个人偏见。会不会早起者 Kripke 在傍晚驾驶时感觉最受损害,而猫头鹰 Stickgold 在早晨感觉不佳?我瞥了一眼统计数据,注意到事故率压倒性地遵循人类昼夜节律睡眠倾向的平均。这意味着两位博士都有道理。可能正是睡眠时型决定了不同时段事故机会的平均差异。驾驶时的昼夜节律相位是更好的决定因素。我的猜测是,任何损害清醒日回忆或记忆巩固的因素也会损害驾驶。换言之,在 SuperMemo 中绘制的警觉性图可以轻易用于确定一天中任何时间驾驶时交通事故的概率。在调查不同年龄的事故率时出现了有趣的差异。中年人在早上 6-7 点有巨大的事故激增,这与 Kripke 的主张矛盾。然而,对于较年轻的年龄,整个夜间的事故率都很高。对于不需要早起、也不需要疯狂熬夜驾驶的退休者,峰值与他们的午睡时间对齐。无论真相如何,Kripke 博士用他的陈述为短睡眠辩护时绝对是错的。根据国家睡眠基金会,每晚睡眠少于 5 小时的人发生车祸的可能性是睡 8 小时或以上者的 4 倍,即便睡 6-7 小时的人风险也翻倍。显然,如全文所论证的,重要的不是睡眠长度而是质量。那些在自由睡眠中获得 5 小时的人不必担心。
我自己的偏见(My own prejudice)
早起者很难理解夜晚型人的问题,他们被自己的状况所偏见。科学家常常分裂成激烈对立的阵营,这些阵营常基于来自琐碎来源的严重偏见,这很好。发现受益于激情,即便它们常常且不可避免地跨越理性边界。我也严重偏见,我敢于批评的那些杰出科学家也是如此。我打赌当 Horne 或 Siegel 博士读到 Stickgold 或 Walker 阵营的新想法时,他们可能自言自语”哦不!又来了……”如果他们读到本文,他们可能惊呼”那个该死的 Wozniak 是谁!?“那是健康的,如果他们愿意读,我会感到荣幸。
我打赌一些自己是短睡眠者或早起者的研究者,倾向于从自己的位置推断到更广泛的人口。看过数百份 SleepChart 日志,我知道一些人确实需要 9-10 小时睡眠,得不到就感觉糟糕。可能有潜在的健康问题吗?也许。然而,其中大多数是其他方面相当健康且是好学生的青少年。我也看到过只需每晚 4 小时、偶尔只需 2-3 小时就感觉极好的人的日志!两个极端都是少数。大多数人在自由睡眠作息下 6.5-7.5 小时过得很好。
当站在闹钟和短睡眠对立面时,我必须划定我自己的偏见。我一生中很少使用闹钟,12 年前我决定彻底摆脱闹钟。我完全爱上了我不间断的睡眠,想与每个人分享这份乐趣,尤其是年轻一代。与 Horne 博士的主张相反,尽管我自由运行睡眠,我相当是个短睡眠者。当我写这些字时,我最近的夜间睡眠长度是 5.7 小时,远低于人口平均。自从我十多年前开始严格自由运行睡眠以来,它一直在减少。自从我近 20 年前开始双相睡眠以来,它减少得更多(见:napping 对夜间睡眠和总睡眠时间的影响)。我从未注意到仅仅因为环境有利(长夜、冷天、雨天等)就睡得长的倾向,恰恰相反,我越虔诚地遵守好睡眠规则,我的睡眠越短,学习质量越好(用 SuperMemo 测量)。尽管这一观察肯定是我自然且不可避免的认知锚定,我在数十份 SleepChart 提交中也注意到同样的相关性。
我对自由睡眠的热爱并不意味着我不知道严重睡眠剥夺的感觉。我记得 1990 年代初在 SuperMemo World,作为程序员赶截止日期工作时,我曾大约 72 小时不睡。由于工作的压力和兴奋,我没有感觉脑死亡。然而,结果近乎灾难:一张装有成品的 CD-R 正在送往德国母带制作和生产的路上,测试中发现了一个灾难性的 bug。那次睡眠剥夺和截止日期本可能非常昂贵。
如果我没有尝试自由睡眠,我可能对 Stickgold 博士的夸张相当怀疑:“睡眠剥夺让你变胖、生病和愚蠢。” 在 1980 年代,我处于与质量睡眠的持续战斗中,每年通宵十几次,几乎总是晚睡,我实际上相当健康,非常瘦(主要因为来自贫困家庭),并以优异成绩毕业。既不胖,也不病,也不可认证地愚蠢。然而,既然我已经尝试自由睡眠十多年,我的观点大大改变了。我现在比 20 年前健康得多,学习也快得多(尽管那种加速大多归功于技术和经验)。我可能仍在为维持健康的体脂水平而日常斗争,然而,那场斗争在 20 年前 SuperMemo World 早期、我的睡眠卫生处于最低点时最为艰难。我坚信自由睡眠改善了我的健康和创造力,并相信正是自由睡眠帮助我消除了感冒和流感的问题(尽管冬泳可能也是强有力的贡献者)。最重要的是,我喜欢几乎忘记了睡眠剥夺的感觉。没有什么比糟糕的夜间睡眠更有效地破坏创造性工作了。鉴于我自己的经验,让我冒昧重写/柔化 Stickgold 博士的主张:
睡眠剥夺会让你更胖、更病、更笨!
Robert Stickgold 博士,哈佛医学院精神病学副教授(意译)
我承认!我严重偏见!尝试过好睡眠后,我根本无法考虑睡眠限制或人为睡眠控制。然而你不必信任我的判断,如果不确定,就应用帕斯卡赌注,把睡眠当作上帝,你的智力正受到威胁!
好睡眠是通往美好生活宝库的钥匙,不要让任何人抢走!
睡眠时长和睡眠相位对学习的影响(Effects of sleep duration and sleep phase on learning)
我在 2000 年代初开始研究睡眠对学习的影响,源于简单直觉:短夜间睡眠对第二天的学习不利。用 SleepChart 收集两年数据后,我试图展示睡眠长度与学习质量之间的联系。然而那次尝试不成功,我的问题在于使用了自己的睡眠和学习数据。我是自由睡眠的虔诚遵守者(即禁止所有形式睡眠控制,尤其是闹钟的睡眠)。仔细审视后发现,在自由睡眠中,短夜间睡眠常常是命中最佳睡眠相位的指示,而长睡眠可能源于过早入睡、繁重运动、健康不佳和其他因素。解读类似于 Kripke 博士的研究(显示睡得少的人活得更长)。那项研究让许多人得出保持短睡眠健康的错误结论。在 Kripke 和我自己的研究中,混淆来自这样一个事实:自然短夜间睡眠是良好健康、正确睡眠相位或长寿前景的指示。使用他人的数据,我后来能够表明,由闹钟使用造成的短睡眠对学习有负面影响。可以预期闹钟对长寿有同样的影响。现在显而易见,在自由睡眠中,睡眠长度不能用作睡眠质量的指示。
在自由睡眠中,总睡眠时间与学习表现之间几乎没有或没有相关性。这种相关性只在睡眠事件长度被人为控制时出现。
入睡时间与睡眠相位的关系比总睡眠量更重要。我们应始终在生物钟说是主观夜晚开始的时候睡眠。在正确时间(主观夜晚)睡 2-3 小时,比在错误时间(例如在日本有时差时)睡 8 小时更有益。显然,在没有严重先前睡眠剥夺的情况下,在主观白天获得 8 小时相当困难。
在用自己睡眠数据进行初步调查时,我得出结论,我可以用睡眠相位作为睡眠质量的更好指示。SleepChart 使分析数据并近似最佳入睡时间成为可能。那些预测非常初步,可以用以下推理解释:如果你昨天凌晨 5 点入睡,且那个晚是自然的、非强制的,不要指望今天能在凌晨 2 点进入高质量睡眠。在自由睡眠中,疲倦始终是告诉你何时入睡的最终判官。然而,SleepChart 可以警告你疲倦何时可能是内稳态的,是最佳睡眠时间的”不可靠预测”。如果你过早入睡,睡眠会过长、不完全令人神清气爽,并带有过早醒来的风险。如果你过晚入睡,睡眠会不自然地短,并带有移动睡眠相位(即第二天更晚入睡)的风险。这些观察提供了睡眠相位可能影响睡眠质量和次日学习质量的可靠怀疑。这里的睡眠相位是最佳入睡时间(例如由 SleepChart 预测)与实际入睡时间之差。然而,当试图把睡眠相位与学习质量相关联时,我再次失望。我找不到睡眠相位与学习质量(例如 SuperMemo 中的成绩表示)之间的相关性。那次初步努力有两个主要弱点:
- SuperMemo 使用午夜切换概念,允许午夜后执行的复习登记到前一学习日。换言之,在旧版 SuperMemo 中,无法说 5 月 11 日执行的复习是发生在 5 月 11 日还是 5 月 12 日的凌晨。
- SleepChart 过去用简单统计预测最佳睡眠相位,不使用睡眠调节的双过程模型,只是简单的平均值,甚至不用相位响应曲线登记可能影响睡眠相位的区块移动。换言之,SleepChart 只在自由睡眠未被强制延迟、运动、健康问题、阳光等因素干扰时才强。
SuperMemo 和 SleepChart 此后都大大改进。SuperMemo 登记每次复习的时机,SleepChart 依赖相位响应预测昼夜节律顶点。随着数据收集的改进,我能够初步窥视入睡相位与学习表现的关系:
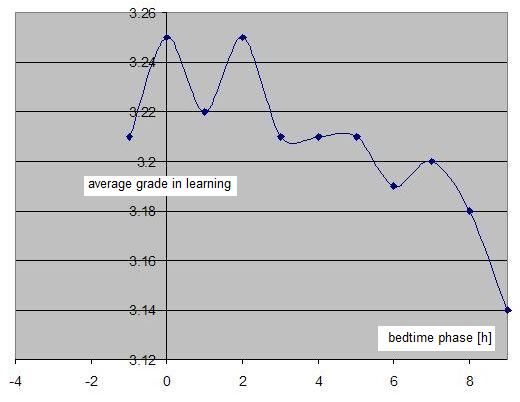
入睡相位与学习表现的关系。入睡相位定义为实际与最佳入睡时间之差。学习表现用 SuperMemo 学习中获得的平均成绩测量。
如预期,推迟睡眠导致表现的逐渐下降。“提前”侧的数据远少,因为在自由睡眠中,过早入睡几乎不会导致早睡,更可能只是在床上一些无效的清醒时间。
在错误相位睡眠(即过早或过晚),会导致学习表现退化。
自 SuperMemo 14.0(2008)起,SleepChart 已成为 SuperMemo 的组成部分。基于这两个应用收集的数据的一些发现列在本文后面的章节。
睡眠区块长度分布(Sleep block length distribution)
NREM-REM 睡眠周期大约 90 分钟。一个流行迷思说,健康夜间睡眠事件的长度因此总是 90 分钟的倍数。另一个迷思说,在 90 分钟的倍数之后打断睡眠是可以的。两个迷思的变体说,睡眠应持续特定于给定个体的周期的倍数。
SleepChart 显示所有睡眠事件长度的分布。该分布可用于证伪睡眠区块聚集在 90 分钟倍数的主张。在所示例子中,单相睡眠者的睡眠区块长度分布确实显示聚集在:1、2、3、3.5、4、5、6、7、8、9 和 10 小时。然而仔细审视,这种聚集仅来自受试者不准确的记录(标记 3.0 小时区块比 2.95 小时区块容易)。没有可辨别的 90 分钟趋势,然而,可能有人被诱惑注意到 60 分钟的倍数。
这是另一个使用半对数刻度的例子,更适合可视化习惯性小睡者的短睡眠区块。在这个案例中,双相睡眠者只在 7 小时睡眠处显示一个显著聚集,这个聚集同样由不精确的记录造成。
最后,一位规律每晚 7 小时的单相睡眠者的睡眠区块分布。可辨别几个峰值,然而没有规律的睡眠长度倍数,尤其是没有在预期的 5.5 和 8.5 小时附近的峰值。
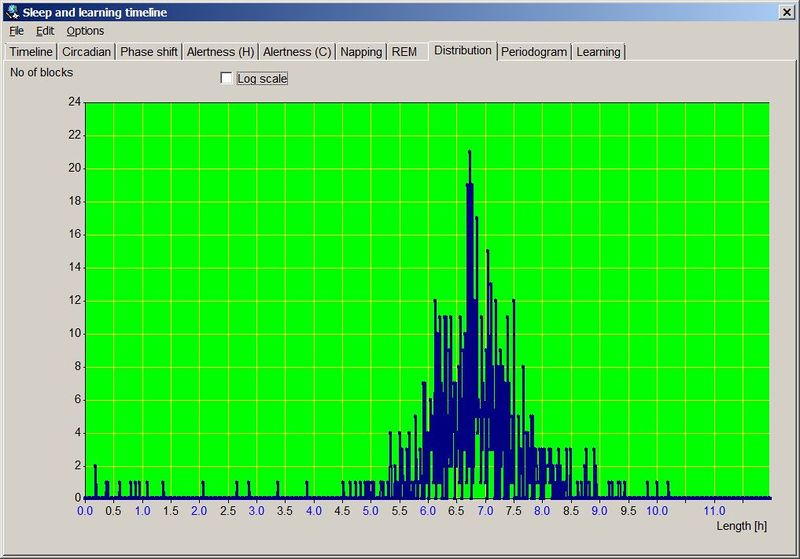
睡眠如何影响学习?(How sleep affects learning?)
为什么睡眠对学习重要?(Why is sleep important for learning?)
如果我要打赌阻碍工业化国家学习的两大因素,它们会是:
- 压力,它带走你的专注、扼杀创造力、榨干动机,并可能促成糟糕睡眠;
- 睡眠,它对优化记忆不可或缺。没有睡眠,你甚至无法体验”美好一天”的感觉。
健康也重要,但从统计上看,正是压力和糟糕睡眠影响着几乎每个人,并造成最大的损失。减少压力、改善睡眠,你可能会看到一个面目全非的社会!
对健康人来说,所有其他学习因素似乎都有些次要。如果你休息好、快乐,自律会大大改善,学习的乐趣随之而来。你接近学习的方式、工具和技术、你在脑海中表征知识的方式等因素,都可以逐步、一致地改进。如果你在稳步前进的道路上,成功几乎有保证。打个比方,你的大脑带着扎实的进步保证,你可以用压力和/或糟糕睡眠轻易使它失效。
鉴于睡眠的重要性,除非你是”天生好眠者”且很少有糟糕的夜间睡眠,否则你应了解睡眠生理学基础和你的睡眠习惯对学习的影响。此外,即便你今天睡得好,你始终面临通过使用电脑、互联网、移动电话、SuperMemo 等毁掉睡眠模式的风险。简而言之,人类大脑还没有足够时间演化和适应现代生活方式的刺激,这就是为什么我们在工业化国家目睹睡眠障碍的流行。
在以下章节中,我将试图表明睡眠对学习的影响远超”休息好的头脑”这一简单概念。
睡眠与学习研究(Sleep and learning research)
每个人都知道,没有一夜好床,第二天可能被毁掉。困倦时,你可以在新鲜空气中轻松铲花园,但如果你在温暖的房间里在电脑前尝试一些创造性工作,你的大脑会倾向于关闭并扼杀任何创造性进展。
认知功能和学习显然是睡眠剥夺的主要受害者,这是很明显的。科学家长期怀疑睡眠的主要功能与学习和记忆相关。甚至在 17 世纪,John Locke 就因这些原因为孩子争取好睡眠。然而,只有近几十年和近年才带来证据的指数级增长,证明睡眠在记忆中的作用。仍有杰出的睡眠研究者质疑这一联系,一些人坚持只有有意识的大脑才能参与记忆,另一些人声称睡眠像进食,如果能获得更多,你总想要更多。在科学界之外,睡眠受到惊人的漠视,许多人不想在睡眠上浪费时间以节省更多时间工作和”创造”,另一些人试图在最短时间内获得”最佳”睡眠(见:Polyphasic sleep)。
那种漠视最糟糕的部分是,全世界的小孩清晨被早早叫醒去学校”学习”。不仅他们的学习受影响甚至变得徒劳;不仅这些孩子变得压力和暴躁;他们的健康可能受影响,免疫系统被削弱,长期发育被阻碍。一些睡眠研究者试图为更合理的学校作息而战(向 Mary Carskadon 博士和 Amy Wolfson 博士致敬;见采访)。同时,无处不在的激烈竞争在美国、欧洲及更远的地方产生了坚持更早学校时段的力量,这些力量来自父母和当局,他们都提出一个虚假且生物学上站不住脚的借口:孩子们可以早点睡。
在这一阴郁黯淡的情景中,仍有一线希望。科学缓慢渗透到社会意识中,然而,最终它大多数时候会赢(除非它需要对抗更强大的力量;例如智能设计理论在宗教教条主义者支持下仍然活得很好)。我乐观地预测,迟早政府、学校当局和父母会意识到,用闹钟把孩子从床上拽起与教育目标相矛盾!
用 SuperMemo 研究睡眠与学习(Studying sleep and learning with SuperMemo)
三十年来,我一直关注现代生活方式对睡眠和学习的负面影响。我建议,很大一部分睡眠障碍可以用时间疗法、自由睡眠等简单技术来治愈。解决睡眠问题的第一步是理解自己的睡眠模式。为此,我鼓励有睡眠问题的人用 2003 年发布的 SleepChart 免费软件收集睡眠数据(下载)。SleepChart 创建时,尚不清楚它会带来什么好处。我建议 SleepChart 未来可用于研究睡眠与学习之间的联系,并可作为优化学习的工具,尤其是与 SuperMemo 结合使用。SuperMemo 的美好之处之一是,它在你学习时保留记忆表现的详细记录。如果该记录能与学习前后睡眠质量的测量结合,将涌现研究机会的海洋。正是 SleepChart 应用提供了缺失的环节。凭借 SuperMemo 和 SleepChart,我们可以收集数据,提供关于睡眠与学习几乎所有问题的答案。然而,我关于整合 SleepChart 和 SuperMemo 的建议遇到了很多反对,主要来自 SuperMemo 用户,他们一直抱怨程序陷入了复杂度不断上升的无尽螺旋,很少有用户会需要或使用新功能。
长睡眠导致糟糕学习?(Long sleep results in poor learning?)
自 1996 年起,SuperMemo 可以保留所有复习的详细记录,你可以查看哪块知识被复习、何时、结果如何。自 2000 年 1 月起,我保留了自己睡眠时机的详细记录,我始终好奇睡眠如何影响学习、学习如何影响睡眠。手头有学习和睡眠数据,我可以寻找两者之间的相关性。我最初最原始的直觉是,应该容易证明短睡眠产生糟糕学习。“更多睡眠是否帮助学习?” 我用自己的睡眠和学习数据快速调查这种相关性。然而,几乎可以证明反向关系。回顾起来,这个悖论很容易解释:在我虔诚实践的自由睡眠中,睡眠质量与长度之间存在相关性——睡眠事件与昼夜节律的对齐越好,睡眠越短、质量越好。除非睡眠剥夺,健康人只有在错误相位睡眠时才会睡得长。最佳睡眠通常非常短。换言之,睡眠长度不是睡眠质量的衡量标准。
学习减少睡眠需求?(Learning reduces the demand for sleep?)
一个类似的问题是*“学习是否增加睡眠需求?”* 当我试图调查这个镜像问题时,我同样不成功。再次可以注意到反向相关。这次惊讶的原因是,睡眠不足会抑制学习。这样,更少睡眠意味着更少学习,以及次夜更长睡眠以偿还睡眠债。换言之,大量学习会矛盾地伴随很少睡眠!更多细节见:Impact of learning on sleep。
这些失败表明,在不考虑昼夜节律时机(即学习相对于睡眠相位发生的时间,例如由自然醒来小时确定)的情况下,几乎无法获得关于睡眠与学习关系的证据。
近似睡眠相位(Approximating the sleep phase)
下一步,我希望看到学习与睡眠时间及睡眠相位之间差距的相关性。然而,要让这种相关性可计算,需要对昼夜节律相位有良好估计。SleepChart 使用粗略的启发式算法试图做到这一点,然而该算法太弱,无法解读由推迟睡眠、压力、力竭运动等引起的睡眠节律重大扰动。该算法在 SleepChart 2.0 中被替换,后者使用递归相位响应曲线(rPRC)估计昼夜节律顶点。rPRC 是相位响应曲线的变体,仅基于睡眠日志记录的昼夜节律外向表达,其唯一的相位移刺激是入睡时间相对于最佳入睡时间的延迟。
复习的时机(Timing of repetitions)
进一步研究的另一个障碍是 SuperMemo 中一个叫做午夜时钟切换(Midnight clock shift)的功能。它使使用昼夜节律时间而非时钟时间记录复习成为可能。例如,如果学生在午夜后继续工作,复习登记到前一天而非新日历日。这可能导致睡眠和学习数据错位整整一天。可悲的是,早期版本的 SuperMemo 只保留复习日期,而非精确时间。这只在 SuperMemo 13(2006)中改变,其中每次复习的时钟时间都被记录。这使得计算每次记忆回忆行为的精确昼夜节律时机成为可能。终于可以把睡眠数据与精确时间框架的学习相关联!我于 2006 年 7 月 17 日获得了 SuperMemo 2006 的 alpha 版本。这意味着,截至撰写时,我们已经过了数据收集的第五个周年,数据集每天都在变大、更有意义。在 SuperMemo 中记录睡眠的人的圈子在扩大。
SleepChart 的影响(The impact of SleepChart)
SleepChart 在 SuperMemo 中的应用在价值上超出了所有预期,现在是研究睡眠与学习的独特工具。截至本文发布(2012 年冬),这是世界上唯一使类似研究成为可能的工具。在这项研究中使用 SuperMemo 是必不可少的,因为它在每次复习时有效地瞄准相同的知识保持水平。这为不同昼夜节律和内稳态睡眠倾向水平之间提供了更稳定的比较平台。其他间隔重复应用的开发者从未表达过对研究睡眠的多少兴趣。此外,最新 SuperMemo 算法 SM-15 的额外准确性提供了额外敏感性,即便对较小数据集也应更快澄清趋势和相关性。
你也可以加入研究努力!在 SuperMemo 15 中,你只需记录睡眠并按一个按钮发送数据。关于 SleepChart 在 SuperMemo 中功能的更多细节可在这里找到。
回忆 vs. 巩固(Recall vs. Consolidation)
在研究睡眠对学习的影响时,我们必须分离两个重要的记忆度量:回忆(recall)和巩固(consolidation)。
回忆测量在任何给定昼夜节律时间可以从记忆中回忆的信息片段比例。在 SuperMemo 中,回忆可以简单地测量为在选定昼夜节律子时段内学习获得的平均成绩。成绩可以转换为百分比回忆,或用作回忆的等效度量。转换可以是全有或全无型(成功回忆视为 100% 回忆,失败视为 0%)。转换也可以依赖 SuperMemo 中的预期和/或估计遗忘指数,提供对回忆难度的更精确反映。使用遗忘指数的转换可以基于成绩与预期遗忘指数的相关性,或使用基于主观估计遗忘指数评估的启发式(注意,估计遗忘指数与预期遗忘指数不同,不是 SuperMemo 中复习历史的一部分)。后者看似不那么精确的方法,在回忆水平之间提供更清晰的对比,通过在 SleepChart 的警觉性图中按下 Exp FI 按钮实现。
巩固测量我们在任何给定昼夜节律时间用复习巩固或重新巩固记忆的程度。回忆测量快速,我们在学习当天获得数据,立即知道我们能否在选定时间回答问题。然而,巩固数据可能需要数年收集。我们今天复习一个项目,可能需要等几年才能验证复习的结果(巩固)。由于 SuperMemo 中的睡眠与学习选项相对较新(复习时机自 2006 年起收集),只有收集多年的非常大的数据集才为有意义的巩固测量提供基础。因此,记忆巩固图目前不是 SuperMemo 数据分析的一部分。计划在几年后当它们对更大比例的长期 SuperMemo 用户可用时引入这些选项。
回忆(Recall)
用 SuperMemo 收集的数据显示,回忆随清醒时间快速下降。
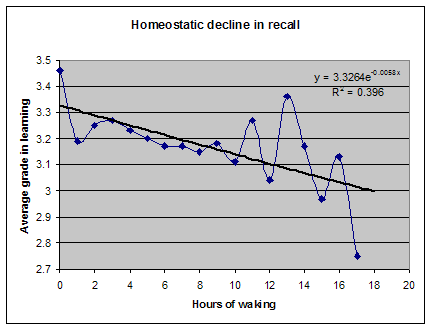
一天中回忆下降速度的示例说明。在本例中,平均成绩从一天早期的 3.3 降到清醒 16 小时后的不到 3.0。
随着一天过去,我们从记忆中回忆事实的能力越来越差。
有趣的是,即便短暂的小睡似乎也把回忆带回基线水平。换言之,回忆和警觉性之间似乎有直接联系。回忆似乎与内稳态睡眠驱力呈负相关。清醒约 12 小时后回忆的轻微增加反映警觉性的昼夜节律组分。图中较晚清醒小时看到的波动来自数据稀缺,因为较晚小时的学习意义不大(用于绘制图的 31,000 次总复习中,只有 684 次落在清醒第 10 小时之后)。
更新版的 SuperMemo 让每个人都能看到自己的昼夜节律周期与回忆之间的关系。
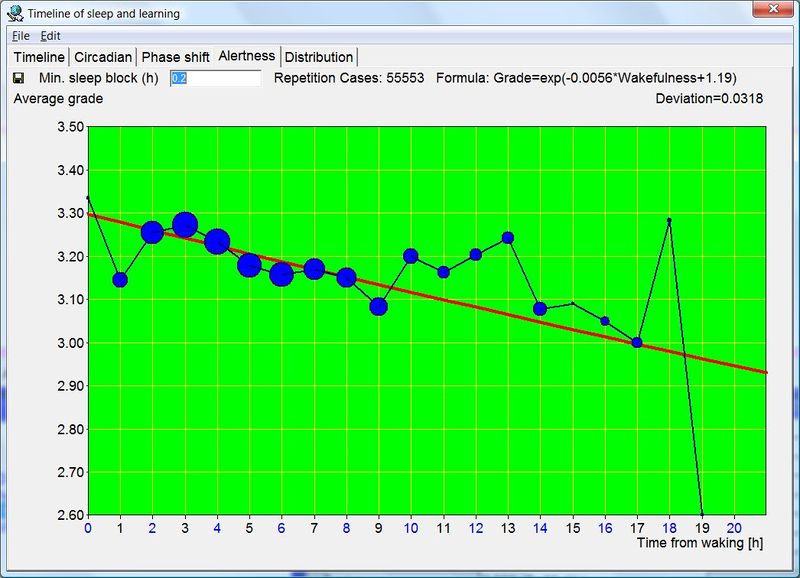
SleepChart 显示的示例回忆图,显示清醒一天中学习评分成绩的下降。该图还显示,由于昼夜节律原因,下半天成绩略有增加。
注意上面两张图都显示类似的时间常数,分别为 178 和 172(半衰期为 124 和 119 小时)。出于校准原因,半衰期只有在使用实际回忆百分比数据时才有意义(在 SuperMemo 中,3.0 是回忆成功与失败之间的尖锐边界)。
记忆巩固(Memory consolidation)
清醒一天中巩固记忆能力的下降,遵循一条镜像从记忆中回忆事物能力下降的曲线!
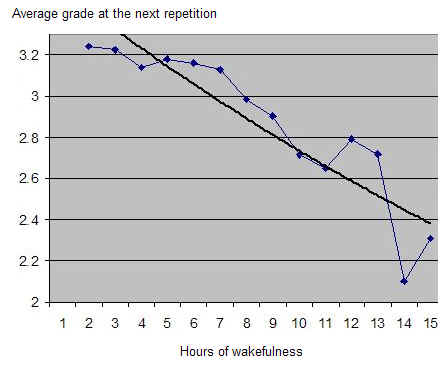
昼夜节律时间(距醒来小时)与记忆巩固能力(用下次复习获得的平均成绩表示)的示例关系。
随着一天过去,把事实存入记忆的能力下降。SuperMemo 中的一次复习是从记忆中回忆先前所学信息的单一努力。该图通过把一次复习的昼夜节律时间(相对于醒来时间)与同一信息片段后续复习获得的成绩相关联而构建。后续复习常常发生在登记巩固时间的复习之后数月或数年。再次,短暂的小睡似乎把记忆巩固能力恢复到基线。与回忆一样,巩固似乎与内稳态睡眠驱力负相关。在自然醒来约 12 小时后也可以看到学习质量的轻微增加(在本案例中)。
结论是,在自由睡眠中(即主要在没有闹钟的情况下),我们如果在清晨学习可以获得最佳学习结果。考试也是如此,如果考试在早晨举行,回忆和考试结果最佳,即便考前临时抱佛脚的时间可能扭曲结果。
回忆与巩固之间的相关性(Correlation between recall and consolidation)
回忆和巩固曲线在清醒一天中似乎遵循非常相似的进程,这一事实表明它们可能都依赖相同的底层机制。这一结论被以下事实放大:回忆是被动过程,而巩固是形成新记忆或重新巩固旧记忆的主动过程。我们可以假设底层机制因此不是分子层面的。回忆和巩固的下降可能简单地由参与学习的神经网络运行效率下降引起。那种效率(表示为警觉性,见:Alertness in SuperMemo)取决于睡眠驱力的内稳态和昼夜节律组分。内稳态组分决定清醒一天中网络效率的总体下降,而昼夜节律组分允许在清醒日下半天有一个小凸起,可能是由于昼夜节律周期对中枢神经系统整体功能的神经激素影响。
好学习日(Good learning days)
回忆与巩固之间的相关性也可以从昼夜节律时间抽象地看到。如果从学习过程的各个日子取总体回忆和巩固数据,它们也相关良好:
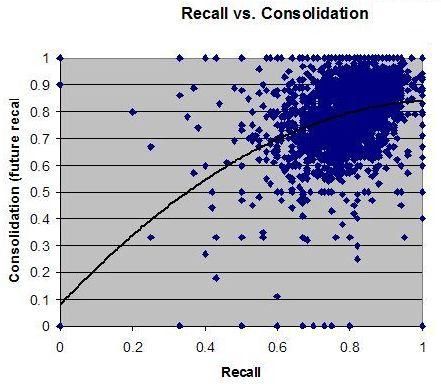
显示对回忆好的学习日也对记忆巩固好的示例图。回忆表示为给定一天正确答案的比例。巩固表示为巩固测量那天的复习之后下次复习那天的正确答案比例。
我们可以得出结论,好的学习日对回忆和巩固同样好。更一般的结论是,成功的回忆对记忆巩固至关重要。
在未来版本的 SuperMemo 中,用户将能看到自己的回忆与记忆巩固之间的严格相关性:
显示良好记忆回忆如何改善记忆巩固的示例图。回忆与巩固之间的关系几乎是线性的。该图使用 SuperMemo 中超过 80 万次复习绘制,其中 53.8 万次为巩固估计提供数据。少于 3000 数据点的回忆水平已从图中省略。Deviation 参数说明线性拟合与数据的匹配程度(偏差越小,拟合越好)。偏差计算为近似值与数据之间差的平方的平均值的平方根。
显示产生给定记忆巩固水平的日子平均回忆的示例图。巩固与回忆之间的关系几乎是线性的。该图使用 SuperMemo 中超过 80 万次复习绘制。少于 3000 数据点的巩固水平已从图中省略。巩固为 100% 时回忆偏低,是因为这一巩固水平被小样本日子过度代表——在这些日子里,仅仅少数项目的幸运满分回忆可能导致完美的巩固读数,却并未真正说明进行巩固复习那一天的回忆情况。足够多这样的案例会使 100% 巩固类别通过本图设定的 3000 数据点异常值上限,并导致一个更接近平均水平的回忆读数。
闹钟 vs. 学习(Alarm clock vs. learning)
迫切需要从以各种方式不尊重健康睡眠的受试者那里收集睡眠数据。进一步研究最有趣的领域是糟糕的睡眠卫生如何影响学习。作为例子,让我们看看一张有趣的图,显示一位常常需要为上学早起(远早于自然醒来时间)的青少年的平均回忆。如果把成绩转换为遗忘指数,我们可以看到这位学生在需要早起的上学日忘记多 53%。这是一个非常初步的样本,不应用于得出深远结论(例如,无课的日子有更多学习发生在较早小时),然而,我希望随着更多数据涌入,我们能切实证明早校时间对学习的灾难性影响。在其他数据集中,也发现较晚醒来(11 点后)常常与较低成绩相关(也许是周末晚”派对”导致更糟睡眠和更晚醒来的结果)。
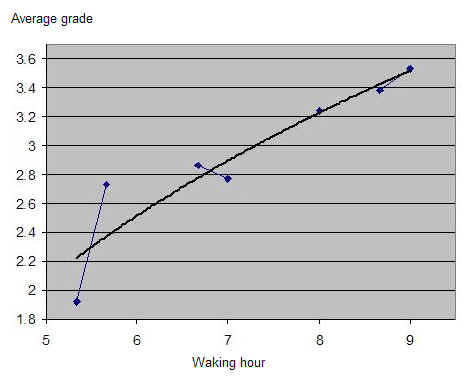
自由睡眠中的学习(Learning in free running sleep)
每个人都有自己的最佳学习时段,取决于昼夜节律。对大多数人,最佳学习发生在早晨和午睡后。不小睡者也因昼夜节律上升而在傍晚改善学习。然而,这些最佳时段的精确时机只能个体化确定。最佳学习时间与绝对时钟之间的脱节,可以在规律的自由睡眠节律中看到,如下面的类似图所示,该图未显示任何学习更高效的时钟小时:
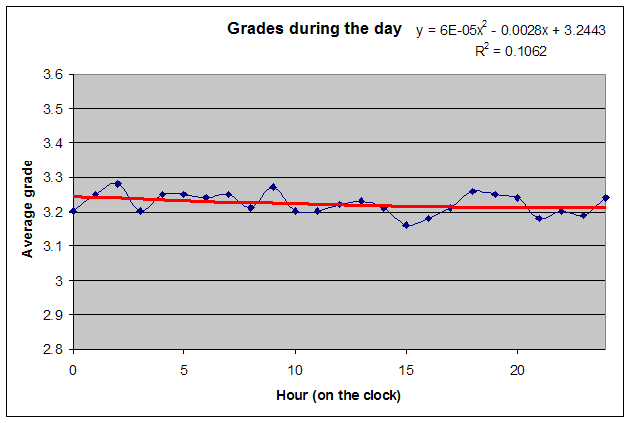
然而,当上面图中呈现的自由睡眠数据用昼夜节律时间而非时钟时间处理时,典型的双峰昼夜节律模式重新出现:早晨好成绩、午睡低谷和傍晚上升。昼夜节律相位估计由 SleepChart 生成。峰值学习时间通常相隔 10-13 小时:
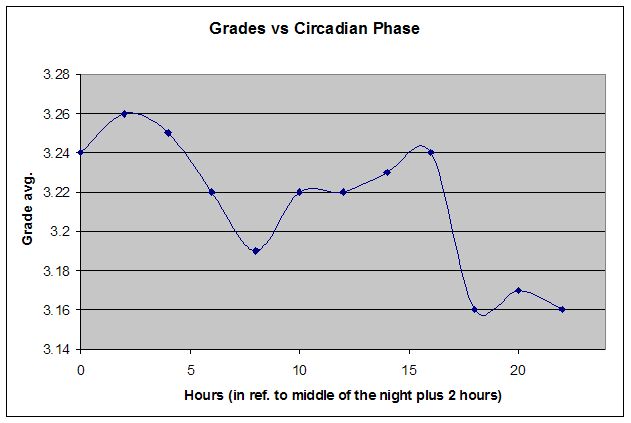
警觉性乘数(Alertness multiplier)
警觉性改善学习是显而易见的。然而,值得注意的是,即便是高警觉性的边际改善也能给学习带来重大收益。换言之,仅仅警觉是不够的,敏锐的警觉性可能比仅仅是”还行”的警觉性大幅改善学习。在所示图中,睡眠倾向用 SleepChart 使用双组分模型估计。
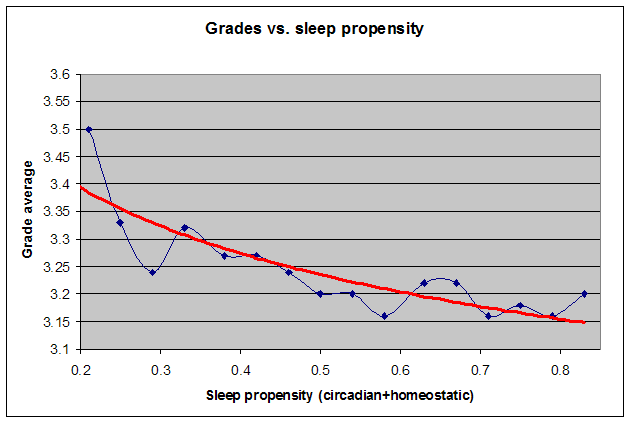
学习过载(Learning overload)
我们在给定的一天花在学习上的时间越多,学习能力越低。回忆沿内稳态困倦增加而下降。然而,当学习过程继续时,它下降得快得多。换言之,学习增加睡眠倾向。这一观察与互补编码理论很好地吻合,这些理论解释了大脑如何应对人工神经网络中发生的灾难性遗忘。这些理论谈到用于重新分配最初存储在低干扰短期网络中的知识的次级记忆系统。存储重新分配的行为被假设发生在睡眠中。换言之,随着你不断向记忆加载知识,你的大脑开启防御机制,让你困倦,并送你更早入睡。这就是为什么我向失眠者推荐 SuperMemo(如果他们必须早睡),这与睡眠专家的传统建议相反。除了缺少困倦的昼夜节律组分的情况外,学习是提升内稳态困倦的好工具。显然,它在考前学习等情况不起作用,那可能潜意识地与压力相关。
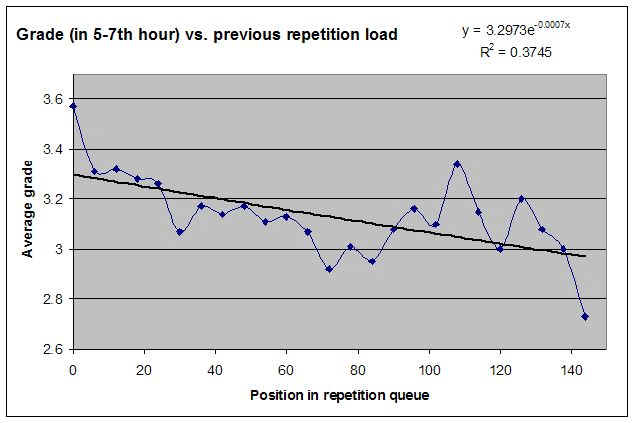
SuperMemo 学习中的平均成绩取决于测试项在学习队列中的位置,靠后的项获得更低成绩。为消除内稳态睡眠倾向的影响,所研究的所有复习都发生在清醒日第 5-7 小时。
睡眠可能是主要的抗过载保护机制。该假设说,睡眠帮助从海马体卸载分离的神经表征,优化长期新皮层的重叠表征。在一夜好睡后用新鲜头脑学习将被推荐。在睡眠剥夺或精神疲劳条件下学习则是错误(除非用作抗失眠策略)。
假设过多学习可能导致阿尔茨海默病的 Robin Clarke 写道(Wozniak,200279):“自然选择将青睐进一步的机制,使接近过载的局部矩阵能够发出缺乏备用容量的信号,从而激活转移到其他位置。” 这听起来正像 NREM-REM 睡眠交互的工作。优化存储是对抗记忆干扰的最简单防御。睡眠可能充当抗过载和抗干扰机制,而不显示与遗忘相同的破坏力。关于”缺乏备用容量”的信号可能简单是双过程睡眠模型中基于腺苷的内稳态组分。另见:Neural optimization in sleep。
警觉性 vs. 学习(Alertness vs. learning)
如前几节所示,在不睡眠剥夺、在正确相位睡眠的健康个体中,最佳学习结果在清晨获得。这一容易复现的观察是 SuperMemo 引入两个选项的激励,帮助用户研究整个学习日的警觉性。在 SuperMemo 中,警觉性一词可互换使用,描述两种不同的认知功能度量:从双组分模型推导的睡眠倾向(或睡眠驱力)的倒数,以及 SuperMemo 学习的平均成绩(对应记忆回忆)。两种警觉性表达密切相关。SuperMemo 测量警觉性,并试图在两个不同的时间框架中预测警觉性变化,旨在分离睡眠倾向的内稳态和昼夜节律组分。两种方法都需要睡眠日志才能进行测量和预测。为展示警觉性的内稳态变化,SuperMemo 测量自上次睡眠事件以来的学习表现。为展示警觉性的昼夜节律变化,SuperMemo 测量相对于昼夜节律时间(即自最佳自然醒来小时测量的时间)的学习表现,时段可能包括或不包括中间睡眠事件。如附图所示,无法完全解卷积内稳态和昼夜节律睡眠倾向对学习的影响。内稳态图总是包括一个与午后小睡学习相关的小昼夜节律凸起,而昼夜节律图将受与昼夜节律周期密切相关的睡眠习惯影响,尤其是在自由睡眠中。
如果你已经用 SleepChart 收集了睡眠数据,你可以用最新 SuperMemo 看到你的清醒-回忆相关性。注意,只有 SuperMemo 13.0(2006)或之后执行的复习才包含在图中,因为早期 SuperMemo 不在复习历史中存储精确复习时间。
你可以通过检查 SuperMemo 中的 Alertness (H) 图,看到你的警觉性、回忆和成绩在一天中下降多快。在该图中,你可以看到自上次睡眠区块以来过去的时间,以及你的回忆在清醒中如何变化:
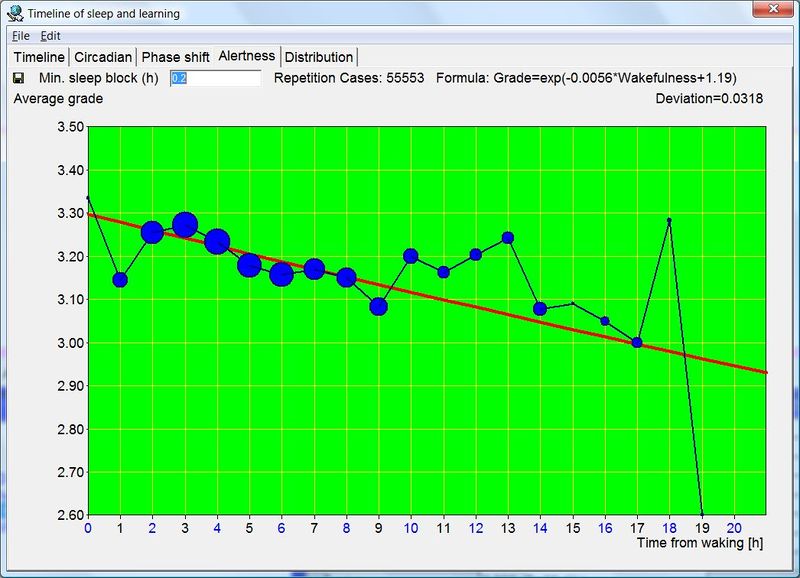
Alertness (H) 图使视觉检查清醒一天中回忆如何下降成为可能。它还显示昼夜节律因素的影响,醒来后立即成绩略低,午后小睡后(即第 10-13 小时段)略高。顶部显示的 Deviation 参数告诉你所选近似曲线与数据的匹配程度(图中:负指数回忆曲线)。偏差越小,拟合越好。偏差计算为差的平方的平均值的平方根(最小二乘法中使用)。
在 Alertness (H) 中,考虑的最小睡眠事件长度由 Min. sleep block (h) 框确定(0.2 小时或 12 分钟是默认最小值)。较短的睡眠区块在绘制此图时被忽略。内稳态警觉性半衰期(小时)告诉你醒来后学习能力何时减半。你可以修改此参数以寻找更适合你情况的曲线(必须按下 Model 按钮)。见 Deviation 评估拟合。该半衰期因人而异,值得注意的是,发作性睡病患者非常短,而天生不小睡者非常长。
警觉性的昼夜节律变化可在 Alertness (C) 图中看到,它绘制一天中相对于从实际醒来时间或最佳自然醒来时间测量的昼夜节律时间的警觉性:
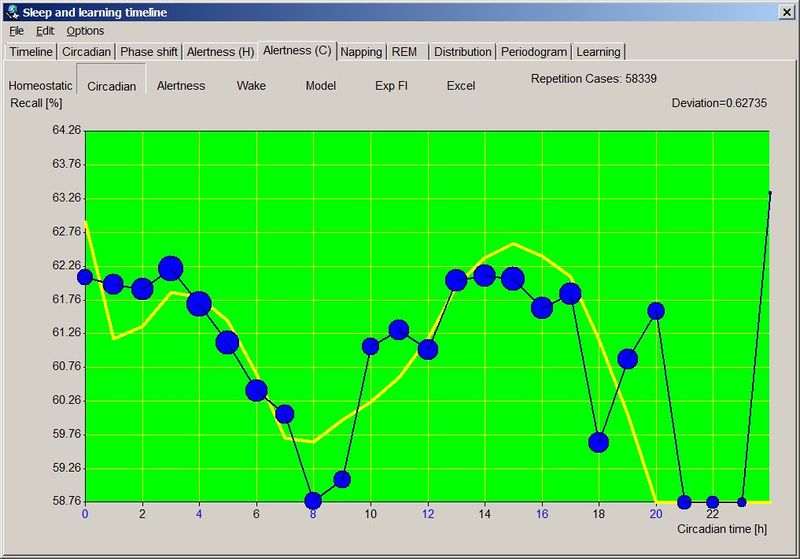
Alertness (C) 图显示人类昼夜节律周期强大的双相本性。横轴显示昼夜节律时间,即从相位 0(预测的”夜晚结束”时间,如果按下 Model)流逝的时间。预测来自 SleepChart 采用的昼夜节律模型,并源自睡眠日志数据。黄线是使用为 SuperMemo 睡眠优化而开发的睡眠倾向双组分模型(受 Alexander A. Borbély 和 Peter Achermann 类似工作启发)从同一睡眠日志数据推导的预测昼夜节律警觉性。图中未显示的总体警觉性是两个睡眠倾向组分状态的合成:内稳态组分和昼夜节律组分。蓝点是从 SuperMemo 学习过程中获取的回忆数据,与总体警觉性相关良好。
学习如何影响睡眠?(How learning affects sleep?)
学习对睡眠的影响(Impact of learning on sleep)
有许多迹象表明,繁重学习增加睡眠需求并增加睡眠密度,尤其是其 REM 阶段(DeKonick,198986、Smith 等,200487)。
在 Learning overload 中,我展示了学习如何抑制进一步学习并如何贡献于内稳态睡眠驱力。在这个意义上,学习确实增加睡眠需求。
在我的许多旧文章中,我常提到学习应增加对总睡眠时间的需求。我在 1980 年代就读到学习对睡眠的影响,此后一直带着这是科学事实(如同睡眠对学习至关重要一样明显)的信念生活。然而,当我试图用 SuperMemo 收集的数据证明这一主张时,我发现这并不像我想的那么容易。
当我试图看先前学习是否增加睡眠长度时,我发现了相反的情况。我再次从自己的睡眠和学习数据开始,这些数据在规模和自由运行条件适用于睡眠和学习两方面都很少见。我在文章前面解释过自由睡眠。所谓”按需学习(learning ad libitum)“,我指的是出于效率考虑,在好学习日更密集、更持久,在较差学习日较不密集的学习。好和坏学习日主要由睡眠质量决定,而非例如时间可用性。我认为自由运行条件对这类研究至关重要,尤其是睡眠不应被人为控制,以确保增加的睡眠需求反映在获得的总睡眠中。
看来,在自由运行条件下,大量学习的日子夜间睡眠更少!
前 8 小时学习量与入睡后前 11 小时获得睡眠量的函数关系。
仔细审视后发现,这一令人惊讶结果的原因是,如果学习按需进行(即好学习日学习更多),先前睡眠质量决定了学习量以及次夜的总睡眠。
在睡眠和学习都按需进行的自由运行条件下,好学习日伴随更少睡眠,因为它们与最小睡眠债相关。
我试图对先前总睡眠进行修正以获得更好的图景。你可能从关于小睡的章节回忆起,小睡量与前夜总睡眠相关良好(睡眠越少,小睡越多)。如果我能找到连续两夜睡眠之间类似的整洁关系,我也许能修正睡眠债,揭示更多学习是否需要更多睡眠。
然而,连续两夜总睡眠之间的关系也相当令人惊讶。例如,下面的 U 型关系显示夜间睡眠量取决于前 20 小时的总睡眠。
总睡眠与前夜睡眠的示例 U 型关系。纵轴上的总睡眠取合并后的夜间睡眠(即忽略短暂夜间醒来的睡眠)。横轴代表总睡眠,其终止点被所考虑入睡时间前 20 小时的边界所包含。选择这个边界是为了捕获前夜睡眠以及随后的午睡小睡,而不触及应被视为距离感兴趣时段两夜的睡眠区域。
U 型图表明,不能用简单的睡眠债公式来修正学习后的睡眠需求。然而,可以使用正常长度夜的子集来过滤掉不同的睡眠债条件。
至于获得的 U 型解释,可能是三个主要原因的组合:
- 最明显的是,一夜短睡眠常常导致次夜更长睡眠(例如前夜总睡眠 0-3 小时)。
- 一些决定自由睡眠长度的因素可能跨越超过一天(例如健康状况、季节、湿度、阳光可得性等),这些因素会导致连续夜总睡眠之间的正相关(如图中总睡眠 5-8 小时所示)。
- 极长的日子,清醒超过 20 小时,将导致前夜睡眠在图上登记为零。
使用连续两夜睡眠长度关系的数据,我们可以对用于生成第一张学习 vs. 睡眠图的数据应用”带通滤波器”。如果我们通过只选择前夜总睡眠等于或超过 5 小时的数据点来消除短睡眠夜,我们可以扭转下降趋势,产生学习与后续睡眠之间近乎平坦的线性关系:
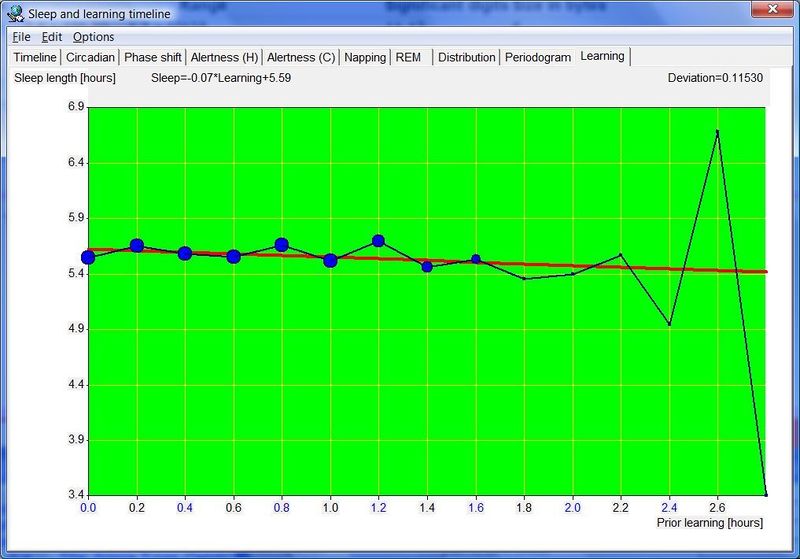
如果前夜睡眠超过 5 小时,则学习量对后续睡眠几乎没有影响。
如果”带宽”缩小到 5.0-6.5 小时,我们得到一条完美平坦的线(斜率=0.00)。这一数据似乎表明,学习增加不增加次夜的总睡眠。
由于有许多证据线表明学习确实影响后续睡眠,这一结论可能有多种解释。睡眠密度可能改变而非夜间睡眠事件长度(如 REM 密度的情况(Smith 等,200487))。在活跃的生活方式中,学习可能不会把睡眠需求增加到远超基线。最后,结果可能因学生而异。一些学生可以发誓更多学习需要更多睡眠。我尚未收到能证明这一事实的适当大数据集。正如自由睡眠无法证明短睡眠对学习有害一样,按需学习可能也无法证明大量学习增加了睡眠需求。正如闹钟可以帮助显示其自身对学习的坏影响一样,强制学习也可能是更感恩的研究对象,强制学习对大脑可能更昂贵,并对睡眠密度和长度显示更明显的影响。也许学习需要足够繁重才能注意到效果,因为我们所有的清醒经验都是一种学习形式,即便我们做无聊的重复活动。一个单纯的思考过程(例如回忆一个亲戚)都会在大脑中形成新的记忆痕迹,这些将在睡眠中处理。要让睡眠需求远超基线,学习也必须远超其基线。
睡眠与学校(Sleep and school)
学校让世界变得更好,全球识字率在上升。然而,有一个巨大因素阻碍学校:困倦的孩子!
困倦的孩子学得少!(Sleepy kids learn little!)
现代生活方式导致青少年人群延迟睡眠相位障碍的流行。如今数百万家庭挣扎着让孩子足够早入睡,并让他们及时神清气爽地醒来上学。似乎我们正在全球范围内输掉这场战斗,孩子似乎获得越来越少的质量睡眠!Amy Wolfson 和 Mary Carskadon 博士研究青少年睡眠,他们惊恐地发现,上学期间的睡眠潜伏期对 10 年级生最低,达到惊人的 1.8 分钟29!这个潜伏期低于一个好睡眠者通常在入睡时达到的值!换言之,孩子们在学校比正常个体在夜间更准备好入睡!
在这种睡眠剥夺状态下学习,价值仅比零多一点!
这是一种令人震惊的状况,可能破坏教育的未来以及下一代的身心健康!一些睡眠研究者敲响警钟,另一些寻找疗法。我没有这个问题的处方,这里我只想呼吁父母和学校更多的宽容和理解。我一生都为每个人的更好教育而工作,然而没有睡眠就没有学习。睡眠足够重要,常常优先于教育本身!我的呼吁是:
缺一两节课也比困着去上学好!!!
这一呼吁的前提非常简单。把一个半昏迷的孩子叫醒上学,意味着一天实际上对学习是浪费的,或者字面上从年轻生命的日历上划掉。增加那额外的 2-3 小时睡眠意味着孩子只会缺一两节课,一天中还有许多额外的高效小时!花一小时高效学习远比花 8 小时半昏迷地”熬过课堂”好。这个简单的真理多么不被欣赏让我惊讶!当我与父母交谈时,他们总是用”缺课会有后果”为早醒辩护!绝不能有任何后果!睡眠剥夺使大脑萎缩!睡眠是发育中大脑的基本人权。如果有人以”后果”威胁孩子,你需要对抗那种态度。可悲的是,对许多父母来说,早晨作息的时机由工作和无法绕开的其他义务决定。
过重的学校负担(Excessive school workload)
我最喜欢的记者之一 Fareed Zakaria 在他的 GPS 节目中谈到他对更好教育的处方:“解决方案的一些元素似乎显而易见。作家 Malcolm Gladwell 说,要在任何事上真正出色需要 10,000 小时。这只是另一种表达 Thomas Edison 著名观点的方式:天才是 1% 的灵感加 99% 的汗水。现在,如果我们的孩子比许多其他国家少在学校待两年,他们会在许多领域落后。我们不必走到韩国延长上学日和学年的地步,但我们不能做最少的工作并指望最好的结果。“(来源)。
这种”韩国方案”的问题在于,它没有考虑到好的学习和坏的学习之间的巨大差异。鉴于好老师、好资金、好方法等的短缺,我们也许可以增加学校小时,希望把数量转化为质量。然而,一小时好的自学或一小时定制的一对一辅导,比 10 小时在普通教室的无聊更有价值。也许依赖优秀教师的芬兰学校会显示更好的比率。如果我们只给学校系统加两个因素:(1) 好睡眠和 (2) 间隔重复(spaced repetition),我们可以安全地把学校小时减到每天 1-2 节课,仍然获得更好的结果!
一小时好的自学比 10 小时在普通教室的无聊更有价值!
学校作息与在家上学(School hours and homeschooling)
由于青春期有充分记录的相位移,青少年发现越来越难以通过更早入睡来解决睡眠剥夺问题。早睡可能不会提供更长睡眠,反而可能导致失眠和一系列心因性睡眠和情绪问题。青少年根本无法在设计师定制的早时间入睡,试图强迫他们这样做可能实际上适得其反。即便是轻微的睡眠剥夺也可能比数小时辗转反侧或夜间醒来更好。回到农民的生活方式可以纠正青少年生物钟的问题,然而,这意味着从清晨开始在田间进行数小时体力劳动,坐在学校长椅上做不到。沉重的学校作业本身也促成了晚睡眠相位的生活方式!
当学校试验较晚的上课时间以适应青少年生物钟时,他们获得了更好的学习结果(Wahistrom,200288)。年轻驾驶员在上学或放学路上的交通事故也下降(仅顺时针推迟一小时就降约 25%)。然而,孩子似乎只是倾向于适应并在夜里熬夜更晚。较晚的学校作息是不完美的疗法,尤其是孩子们因睡眠时型而异,每个人都有自己的最佳学习表现窗口。Callan(1998)报告,在高中,只有不到 10% 的孩子喜欢早校时段,而 15% 喜欢傍晚时段。报告的偏好常被昼夜节律周期与清醒期的错位所混淆,这常让夜晚型声称傍晚更适合学习,而在自由睡眠中,同样的孩子会偏好主观早晨时段(“早晨”可能晚到中午)。此外,随着孩子长大,夜晚性的主导变得更明显。
我不知道通用解决方案,然而,所有父母都应考虑在家上学(homeschooling),这可以产生天壤之别。并非每个父母都有资格,也并非每个人都能负担。令人惊讶的是,一些现代进步的国家完全禁止了在家上学,很难相信,但两个采用理性和科学社会解决方案的领导者——德国和瑞典——属于那一组!出于对危险意识形态的恐惧,一些政府阻止回归一个与人类种族一样古老的传统,这个传统可以纠正学校系统的许多弱点:在最充满爱意的人——自己的父母或其他家庭成员——的监督下进行一对一辅导。在家上学使采用最高效的学习方法变得容易:基于激情和好奇心的自定进度、自导向探索,这一理想解决方案解决了学习时间与昼夜节律周期匹配的问题。
儿童的睡眠剥夺(Sleep deprivation in kids)
大多数孩子比他们愿意的更早醒来,这导致睡眠剥夺和一系列负面后果:
- 糟糕学习:削减睡眠严重损害学习。一些学龄儿童群体在坐在教室时显示仅 1-2 分钟的睡眠潜伏期,这是灾难性的!这是许多人在傍晚入睡时都会羡慕的潜伏期!孩子应在睡眠启动近乎不可能的时候学习!在自由睡眠中,那将是清醒日的前 3 小时!在那之后,学习受损,体育课和午餐可以上场。许多孩子在课堂上睡觉,或全神贯注于”熬过”课堂。
- 糟糕情绪:睡眠剥夺导致易怒。没有睡眠中执行的神经网络清理,大脑很快过载并被刺激淹没,这可以是学习刺激,也可以是同事玩笑等小烦恼。即便在原本有礼貌的孩子中,坏脾气也随之而来。同志情谊和社交互动被欺凌、打架、攻击性和纯粹的恶意取代。多年下来,这可能导致心理问题、抑郁、攻击性、自杀,以及与社会其余部分和谐融合的能力丧失。
- 憎恨学校:到 10 岁,大多数孩子普遍憎恨学校!我遇到的任何孩子,我问的第一个问题就是他/她对学校和学习的喜爱。如果他们声称喜欢学校,常常是因为有机会与朋友互动,或简单地摆脱父母监督或离开家庭环境。假期倒计时成为日常心事,这对学校教育的效率和上大学等长期选择有灾难性影响。普遍的憎恨学校塑造了一代人以及社会应对现代世界挑战的方式,而这一切都始于那个恶性装置:闹钟!
上学日的糟糕回忆(Poor recall on schooldays)
为了说明学校作息对学习的影响,见以下示例图。一位 16 岁高中生在 SleepChart 中记录睡眠模式,在 SuperMemo 中记录学习结果。结合两者,我们可以看到醒来时间与 SuperMemo 学习平均成绩的关系。上学的醒来时间总是早于自然醒来时间,青少年通过周末睡得更长来补偿:
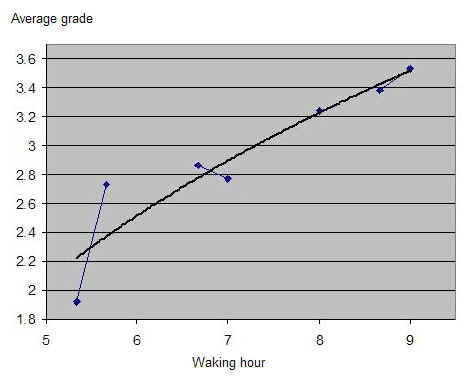
尽管上学日学习表现下降,这位青少年在学校表现出色,在周末进行最佳学习,后来被常春藤名校录取。当成绩转换为遗忘指数时,可以看到睡眠剥夺对学习的戏剧性影响。在这个案例中,学生在需要早起的上学日忘记多 53%。显然,睡眠剥夺不太可能剥夺某人进入常春藤的机会,但它确实影响表现并削弱一个年轻人的潜力。在较小年龄,它还可能对大脑发育有重大影响。有趣的是,在其他数据集中,我也发现较晚醒来(11 点后)常常与较低成绩相关,也许是周末晚”派对”导致更糟睡眠和更晚醒来的结果?
例子(Examples)
例子 1:周末长睡眠(Example #1: Long weekend sleep)
具有短工作日睡眠和长周末睡眠的典型睡眠模式,见以下睡眠日志和相应的昼夜节律图。
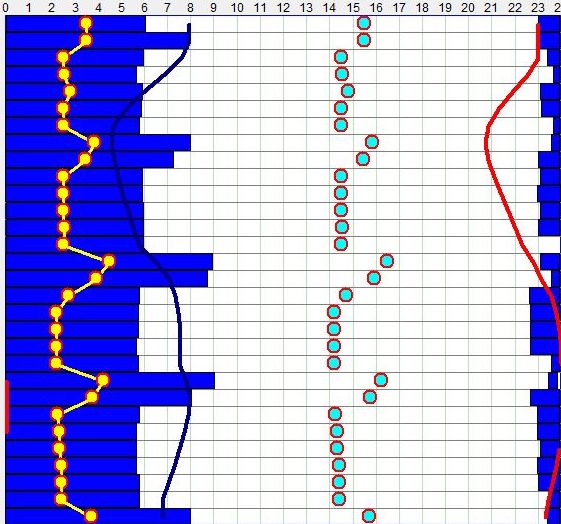
具有工作日睡眠亏欠和周末更长睡眠的示例睡眠日志。通常,周六早晨睡眠比周日早晨睡眠长。
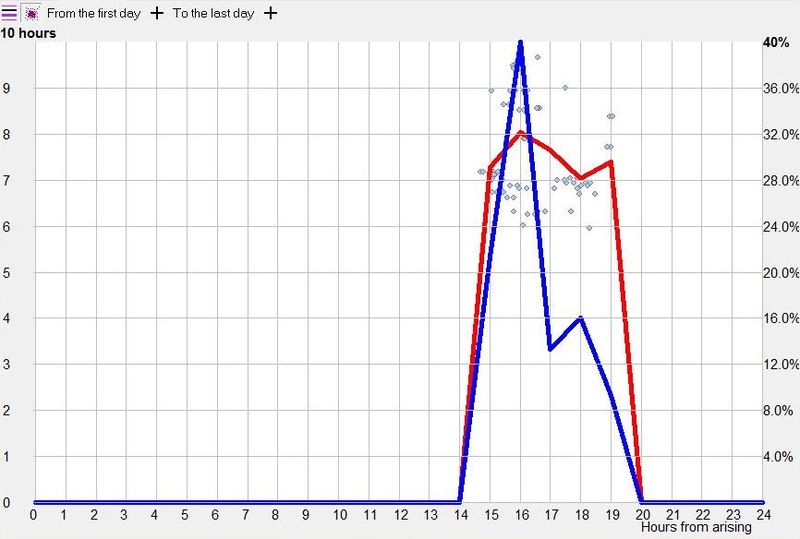
工作日睡眠亏欠和周末更长睡眠的昼夜节律图。该图显示,16 小时清醒加 8 小时睡眠的一天可能是期望的最佳。然而,7 小时的夜间睡眠导致睡眠亏欠累积,工作日每天少睡一小时。
例子 2:相位 12 小睡(Example #2: Phase 12 napping)
一个更令人困扰的例子显示了由短夜间睡眠事件和频繁相位 12 小睡引起的睡眠作息碎片化。这里一位学生试图在完全相同的时段入睡,即 23:00 - 6:00:
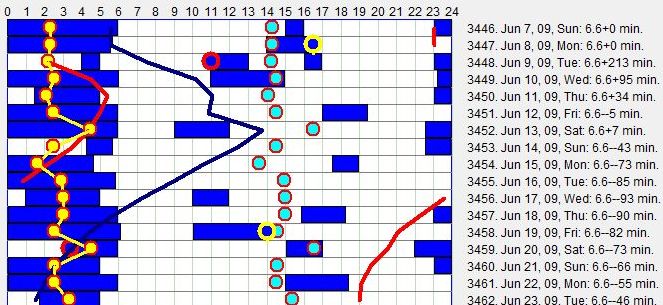
使用不规律小睡补偿睡眠亏欠的示例睡眠日志。
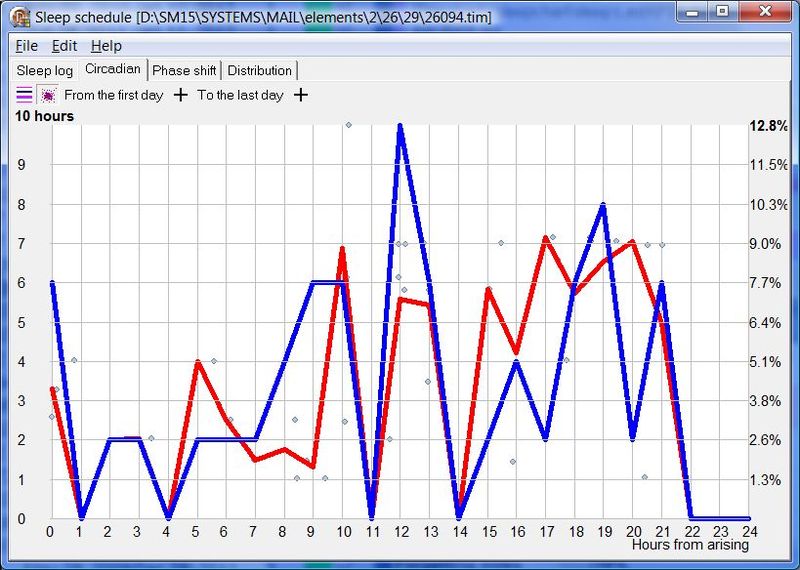
短夜间睡眠伴随不规律小睡的昼夜节律图。小睡在不同相位临时进行。早小睡短,不弥补睡眠亏欠。晚小睡导致夜间睡眠延迟,可能还有加重睡眠亏欠问题的相位延迟。
Alpha 状态下的学习(Learning in alpha state)
有一些学习小玩意和装置被营销为基于放松状态下的学习。恰当的认知环境对学习至关重要。然而,为了清晰,我们应使用”专注(concentration)“一词而非包罗万象的”放松”。学习中的专注应通过考虑以下因素来最大化:
- 切断所有学习干扰源(电话、电子邮件、对话、收音机,也许甚至你最爱的音乐)。
- 找到最佳昼夜节律学习时机(例如自由睡眠周期中的清晨,负担不起自由睡眠的 DSPS 个体的傍晚等)。
- 心理和认知健康的所有方面(例如避免压力、物质滥用等)。
放松的概念常与 alpha 波学习相关,这吸引了许多公司,它们更感兴趣的是底线而非客户实际的学习成功。脑电图测量可以粗略确定大脑当前状态,就像你可以通过扫描周围电磁场来检测大城市的繁忙活动。alpha 波扫描对学习的有用性,可以比作电磁场扫描对城市社交生活的有用性。你需要专注于原因而非症状。alpha 波主要在缺乏视觉处理和其他强烈心理过程时出现,这就是为什么它们不能教条地被视为期望的学习状态。毕竟,入睡前的困倦 alpha 状态正是一天中最糟糕的学习时刻。
在评估”放松产品”时,你需要区分放松效果和实际学习效果。这一领域虚假声明公司数量惊人。很容易陷入学习问题的简单解决方案(例如获得 10Hz 双耳节拍(binaural beats) 差,你的学习问题将终生消失,也许同时你的性欲会改善,你会睡得更好,看起来更年轻)。简单的学习解决方案解释了为什么与”放松中学习”相关的虚假声明如此难以根除。
同时,如果你需要应对压力或失眠,该领域的许多产品可能有合法应用。波兰 Sita 系统的客户开玩笑说,如果公司把产品作为小睡系统营销会更好,这本身就是有价值的应用。1990 年代,我呼吁 SuperMemo 用户让我知道值得一提的、对学习有效的放松产品。我认为至今没有收到任何可信的建议。
睡眠中学习(Learning during sleep)
当苏联研究者声称睡眠辅助教学时,他们启动了一个强大的模因,永远无法重现,现在相当难以根除。你可能听说过提供毫不费力的睡眠中学习的睡眠磁带,它们是苏联声明的直接延续,只是整个睡眠中学习产品系列的一部分。你在睡眠中学习磁带上的投资不会是钱花得值。应劝阻睡眠中学习的尝试! 偶尔可以回忆起睡眠中呈现的一小部分材料,信息可能在短暂醒来或从 REM 到浅睡的过渡期间到达并登记在记忆中。也有充分证据表明,大脑中的一些回路可以在 REM 睡眠中被条件化。然而,睡眠中感官与大脑之间的联系更多聚焦于危险时醒来,而非处理复杂信息。
你从睡眠磁带中可能获得的任何东西,都将远远被对睡眠质量的损害所抵消。如果学习刺激未达到某个阈值,它们将被忽略;然而,超过某个值,它们可能阻止 NREM 睡眠向第 3 和 4 阶段进展,还可能缩短 REM 睡眠。
有趣的是,入睡前几分钟获得的记忆不会被巩固!即便是几分钟的睡眠也会留下一个完全从记忆中抹去的短暂清醒窗口。幸运的是,我们很少在打盹前学习关键任务信息。
不推荐睡眠中学习,并不意味着应劝阻开着电视或收音机入睡。如果你想在入睡前来一剂教育,确保你的磁带、电视或收音机满足这些条件:
- 它们不超过 10-20 分钟自动关闭。
- 它们没有能力把你从时机恰当的睡眠中唤醒。如果你在入睡最初几分钟醒来,可能经历内稳态困倦的剧烈下降,延迟入睡。醒来也可能表明你相对于昼夜节律周期入睡过早。
- 它们不包括高度情绪化内容、令人痛苦的消息、门铃、电话、定时器和闹钟等尖锐声音,因为这些都经过最有效设计以干扰睡眠、打断睡眠。
此外,如果你因一天的压力而难以入睡,微妙的新闻频道可能实际上通过让你的心思远离可能触发 ACTH、皮质醇、儿茶酚胺或其他警觉激素释放的想法来帮助你入睡。
早晨的电视、收音机或磁带也可以,条件是你手动打开(即它们不应作为闹钟替代品)。如果你在预期醒来时间前略早醒来,打开新闻并留在床上,测试你的大脑是否有困倦迹象,偶尔你可能仍能入睡并完成一个有益于智力表现的睡眠周期。
清醒梦(Lucid dreaming)
一些自助个人力量大师不断提起清醒梦作为增强学习 and 创造力的工具。超意识或超现实等术语被抛来抛去。多相睡眠者常声称 Uberman 睡眠作息帮助他们实现清醒梦和增强的现实体验。这一声明可能有一丝真相,阅读多相过山车来理解为什么多相睡眠者可能经历看似更高的亢奋高点(由于完全行尸走肉化时段而显得更高)。清醒梦对学习和创造力的有用性就像 LSD。追求清醒梦更可能扰乱健康睡眠并对学习产生负面影响。在 REM 睡眠中,前额叶皮层通常应被去激活。Hobson 的 AIM 3D 睡眠-清醒空间模型(Hobson 等,200089)可以用来把清醒梦对应的状态说明为一种分区,其中皮层和大脑其余部分占据 AIM 空间中的不同点。这种分区可能干扰 REM 睡眠的生理功能,可以比作慢跑时吃午餐(即向神经系统输入矛盾目标)。使用自动暗示技巧改变 AIM 状态可能影响睡眠中发生的神经过程,后果不可预测且不太可能正面。至于创造力,可以想象 LSD(清醒梦较少)可能提升非特定创造力或帮助理解创造过程。然而,人类大多数创造性突破发生在健康神清气爽的头脑专注于解决特定问题时。幻觉的迷雾无助于把创造力引向有用目的。创造力是概率游戏,你应寻找有意识地引导创造过程的方法,而非不加区分地增加其随机性(Wozniak,200190)。
睡眠的生理学(Physiology of sleep)
我们为什么会入睡?(Why do we fall asleep?)
睡眠的启动(Initiation of sleep)
当大脑产生两个信号时我们入睡:
- (H) “清醒太多了”信号,以及
- (C) “到了通常的睡眠时间”信号。
“清醒太多了”信号被称为内稳态信号(homeostatic signal),“该睡觉了”信号被称为昼夜节律信号(circadian signal)。内稳态信号是网络”疲倦”的反映,你学得越多、想得越多、处理信息越多,精神上就越疲倦,这产生内稳态睡眠倾向。然而,内稳态困倦不足以入睡,你可能因清醒或学习过多而累得要死,却仍无法入睡。这就是昼夜节律困倦出场的地方。昼夜节律困倦在主观夜晚时段最大,还有中午警觉性低谷(也是昼夜节律性的)。当你在内稳态和昼夜节律意义上都困倦时,你终于可以入睡。
内稳态信号(Homeostatic signal)
内稳态信号在大脑的神经网络中产生,与脑电图(EEG)中的慢波活动相关,其已知表达之一是腺苷水平增加,咖啡因阻断腺苷的效应,这就是为什么咖啡可以暂时帮助克服困倦的内稳态组分。同时,咖啡因对昼夜节律组分完全无效,这就是为什么在主观夜晚喝咖啡是不明智且不健康的。随着清醒小时流逝,大脑糖原和 ATP 储备耗尽,ATP 降解为 ADP、然后 AMP、最后为腺苷。腺苷随后在大脑中累积,包括在基底前脑(Porkka-Heiskanen,199991),这是神经内稳态信号的假设来源。糖原储备的耗尽也被假设对内稳态睡眠倾向有自己的贡献(Kong 等,200292)。基底前脑是胆碱能结构,活跃时有助于清醒和 REM 睡眠,其去激活有助于启动 NREM 睡眠和总体睡眠。
昼夜节律信号(Circadian signal)
昼夜节律信号的主要来源是视交叉上核(SCN)。一组基因在调控环路中表达,保持 24 小时的活动节律。SCN 节律可以被光、活动或其他信号重置(见:相位响应曲线(PRC))。SCN 把大部分纤维发送到室旁下区(SPZ)和背内侧下丘脑核(DMH)。SCN 振荡效应产生的激素信号之一,是在主观夜晚从松果体释放褪黑素,这让研究者想到,褪黑素可能是启动睡眠的天然帮手(前提是有足够的内稳态困倦)。
整合内稳态和昼夜节律信号(Integrating homeostatic and circadian signals)
内稳态信号需要与昼夜节律输入整合,整合的精确机制尚不清楚,但有几个可靠的假设。下丘脑前部是假定的整合地点,假设的整合核团包括:内侧视前区(MPA)、前室旁丘脑核(aPVN)和背内侧下丘脑核(DMH)。DMH 和 MPA 向腹外侧视前核(VLPO,负责启动睡眠的主要脑核之一)方向发送大量纤维。
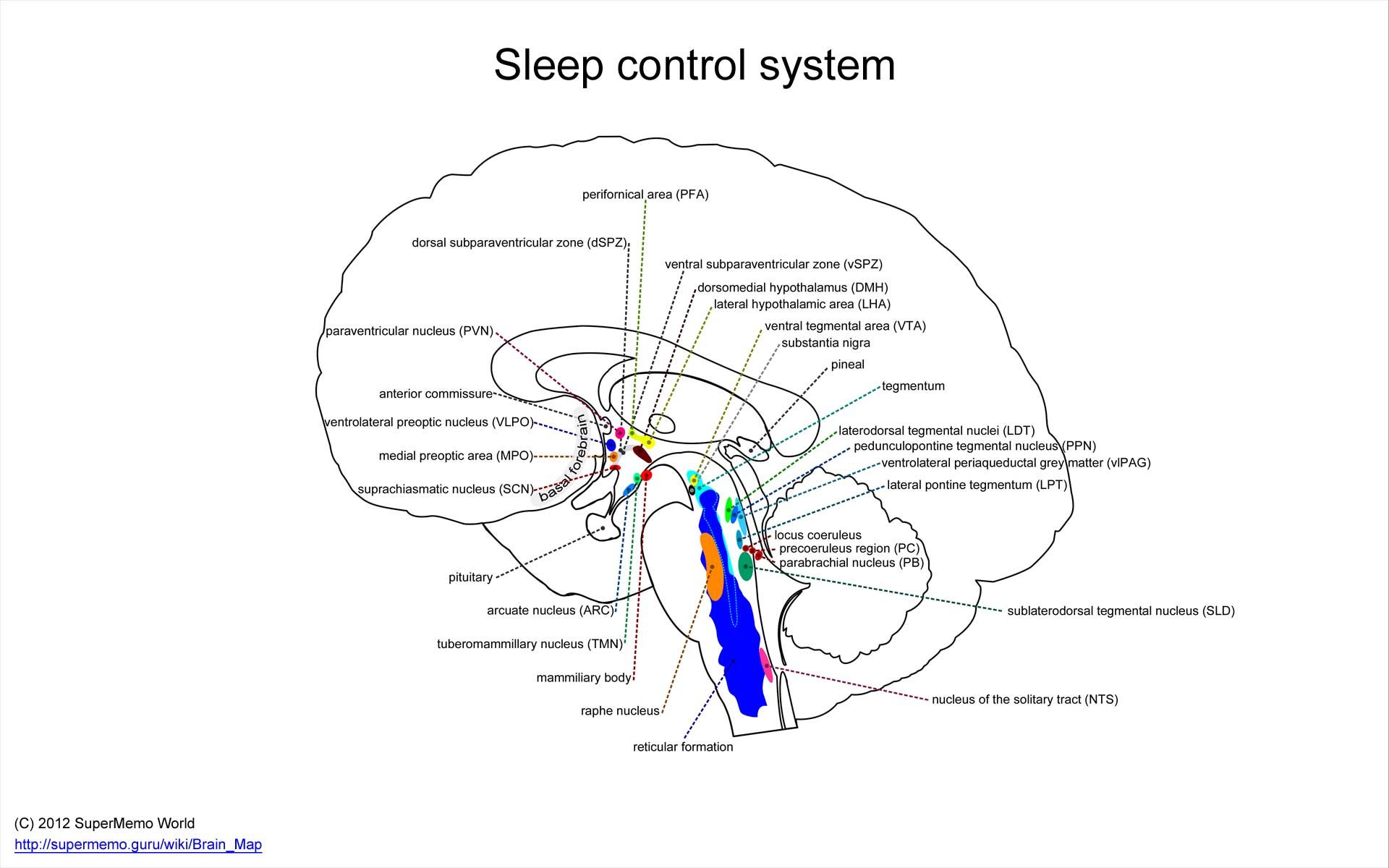
睡眠控制系统。详见正文。
腺苷激动剂也能激活 VLPO(Scammell 等,200193)。有人假设腺苷抑制了下丘脑前部和基底前脑中抑制 VLPO 活动的 GABA 能神经元。
因此,VLPO 通过接收来自下丘脑前部的昼夜节律信号,以及在清醒日中累积的内源性物质(如腺苷)的内稳态信号,能够启动睡眠。VLPO 及其相邻核团随后能抑制促醒的组胺能 TMN 和其他脑桥/脑干唤醒系统(如 LC、DR、LDT、PPT、PeF、vPAG 等)。睡眠是抑制上行网状激活系统(RAS)的直接后果,RAS 是那些保持大脑皮层清醒状态的神经结构的集合。随着 RAS 活动的抑制,我们迅速失去对要求高的智力活动的兴趣,很快我们唯一能想到的就是睡眠。一旦我们在不受打扰的地方休息,我们就飘入梦乡。无法遵循自然身体节律的人,常常无法遵循上述情景。
唤醒的神经抑制还伴随 ACTH 和皮质醇(主要警觉激素)的显著下降,5-羟色胺和儿茶酚胺水平下降,体温也下降。所有这些过程在平行轨道上进行,当它们完美同步时我们睡得最好。通过各种干预(睡前兴奋(多巴胺)、睡前咖啡(内稳态)、运动(肾上腺素)等)把那场交响乐弄出同步非常容易。协同操纵也有副作用:安眠药、酒精或大麻破坏睡眠结构,连褪黑素也有副作用。当所有生理变量按预设的同步变化时,睡眠最健康,这最好通过遵循自己生物钟的戒律来实现。
并非所有科学家都同意(Not all scientists agree)
James M Krueger 博士多年来倡导一个想法:所有高级神经网络都有进入睡眠状态的固有能力(尤其是皮层柱)。Krueger 是受过教育的、有精神气质的生化学家,他从寻找诱导睡眠的物质开始研究,灵感来自一个世纪前的发现:困倦动物的脑脊液含有能转移到其他警觉动物身上诱导睡眠的物质。在过去四十年里,Krueger 积累了大量证据,证明存在大量睡眠调节物质(SRS),如腺苷、一氧化氮、TNF、IL-1、GHRH、前列腺素 D2等(Krueger 等,199994、200195)。一些 SRS(如腺苷)随脑力活动累积(例如谷氨酸释放的结果(Simasko 等,200596)),可能在睡眠稳态中起作用,而另一些(如褪黑素)是昼夜节律的。在近期出版物中,Krueger 断言睡眠是网络涌现现象,大脑中的睡眠控制核团只扮演附属的同步角色。尽管总体原则似乎与神经睡眠控制的主流科学相争,但无可争议事实的主体远大于分歧领域。尽管 Krueger 的理论似乎没有解释睡眠的计算方面(在那里睡眠中心的神经控制似乎不可或缺),但它们都与睡眠控制的内稳态方面相当一致。在生化学家与神经网络专家和神经生理学家相遇的地方,我们可以找到进一步探索睡眠奥秘最肥沃的领域。
昼夜节律周期(Circadian cycle)
人体生物钟以约 24 小时的周期运行,因此被称为昼夜节律周期。理解昼夜节律周期对健康睡眠至关重要。我的粗略猜测是,工业化国家 95% 的睡眠问题源于对昼夜节律周期缺乏理解,或对其力量和重要性缺乏尊重。该周期深深编码在人类基因组中,不易改变或覆盖,玩弄昼夜节律周期可能导致长期健康后果。身体所有细胞都表达各种生物钟基因,然而,大脑中有一个主时钟,帮助同步身体其他时钟,以对健康、福祉、长寿、学习、创造力等至关重要的和谐同步运行。主时钟位于大脑中,称为视交叉上核(SCN)。SCN 的昼夜节律周期导致松果体周期性释放褪黑素,这导致了褪黑素作为睡眠疗法的使用。褪黑素的流行来自其天然来源和口服可能性。然而,由于褪黑素位于昼夜节律级联中 SCN 的下游,它没有产生完整夜间昼夜节律状态的完整魔力。即便褪黑素的自然释放周期,在不规律作息中也可能与睡眠-清醒周期错位,这限制了褪黑素的应用,它可以用来产生相位移(例如在自然入睡时间前 1-2 小时服用产生相位前移),但它不是通常宣传的通用安眠药。
重要的警觉激素皮质醇可用于映射时机恰当的昼夜节律周期,其水平在睡眠前半段下降,醒来时急剧上升,给我们敏锐的清醒头脑。另一方面,生长激素较少依赖时钟,主要在深睡中释放,参与睡眠的合成代谢力量,对大脑和肌肉都很重要。
昼夜节律警觉性部分是激素性的,部分是神经性的。脑干包含一组称为网状激活系统的核团,这些核团激活时让我们保持清醒和警觉。这些”警觉核团”包括血清素能中缝核、肾上腺素能蓝斑、臂旁核等。对这些区域及其连接的各种损伤可能导致失眠或昏迷。
Borbély 模型(Borbély model)
1982 年,匈牙利睡眠研究者 Alexander A. Borbély 发表了一篇题为”A two process model of sleep regulation”的开创性论文6。该模型后来被相当精确的数学术语描述,现在是我们理解睡眠如何启动、以及睡眠-清醒双稳态开关如何在健康睡眠中工作的主流(与实际神经生理学解释抽象开来)。
简而言之,Borbély 注意到了困倦两个组分之间的区别:内稳态困倦和昼夜节律困倦。内稳态困倦在白天由于脑力付出而增加,昼夜节律困倦在夜间增加。Borbély 的模型认为,要获得一夜好睡,你需要在两个困倦组分都处于高挡位时上床。这意味着在昼夜节律困倦到来之前过早入睡是个坏主意,你会不够困而无法入睡,或睡眠浅且易被打断。另一方面,该模型也暗示,过早醒来可能清除内稳态困倦,即便昼夜节律困倦确保我们相当疲倦,我们可能也发现难以重新入睡。
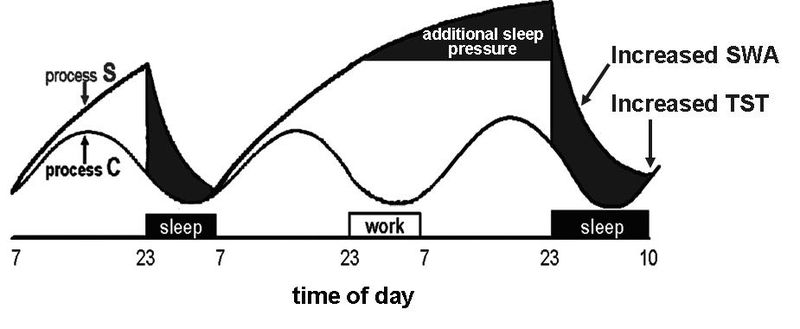
正常睡眠和通宵后睡眠的双过程模型示例解读。内稳态困倦记为过程 S(本文中为助记我用 H),昼夜节律困倦是过程 C 的倒数。当 C 低、S 高时发生睡眠。不眠之夜后累积额外睡眠压力,睡眠可以更早开始并持续更长(尽管昼夜节律困倦略低,它从更高内稳态困倦开始)。SWA——慢波活动——是代表最深睡眠的脑波活动。TST——总睡眠时间——在不眠之夜后更高。
在睡眠中,皮层慢波活动(EEG 功率密度范围 0.7 到 4.5 Hz)取决于先前清醒的持续时间,这就是为什么它被认为是内稳态睡眠倾向的标志(Daan 等,198497),它在入睡后指数下降。该模型的局限之一是不考虑 NREM-REM 交换,而内稳态困倦(过程 S)实际上可能在 REM 阶段增加。昼夜节律困倦与褪黑素释放相关,但也可映射到核心体温或其他睡眠诱导或警觉激素的释放。
Borbély 模型在实践中(Borbély model in practice)
SuperMemo 中有一个受 Borbély 模型启发的睡眠倾向双组分模型,它可以根据睡眠的时机和持续时间预测警觉性变化。
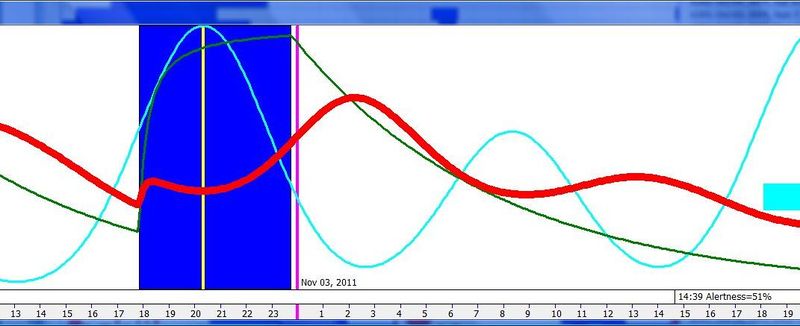
取自 SuperMemo 实际睡眠日志的双过程模型示例性解读。水绿色线代表昼夜节律困倦。绿色线代表内稳态警觉性(内稳态睡眠倾向的倒数)。红线代表总体警觉性,即总体睡眠倾向的倒数。当两个困倦组分都处于最低值时,达到最佳警觉性。
在 Borbély 模型中,睡眠时机由代表两个过程的曲线交叉点决定。SuperMemo 使用更直观的方法,其中两个困倦组分被启发式地整合以匹配预期的总体警觉性进程。当总体警觉性降到某水平以下时启动睡眠。因此,睡眠可以由两个困倦组分独立启动(如在早期生命中),但睡眠的时机和持续时间会因两个变量(和昼夜节律系统状态)的不同值而不同。
你可以”感觉”到两个组分。内稳态困倦更可能被描述为”不神清气爽”的感觉,而昼夜节律困倦更可能被命名为”昏沉”。在健康的周期中,你永远不应看到两者的区别:你醒来神清气爽,傍晚当两个困倦组分都发挥作用让你”非常困”时你变得困倦。然而,如果你有时差且昏沉,你可以感觉到不消失、且在内稳态困倦太少而无法入睡时无法通过小睡缓解的不愉快昼夜节律困倦。另一方面,通宵后你可能累得要死、不神清气爽,然而,随着晨光你获得新能量再熬几小时。当昼夜节律困倦过去,你可能感觉到看似可存活(直到下一个昼夜节律低谷袭来)的内稳态困倦(更多见:Sleeping against your natural rhythm)。
Borbély 模型与演化(Borbély model and evolution)
睡眠的内稳态组分可能只是演化神经网络的不可避免成本。为防止灾难性遗忘,神经网络需要实现过载保护,那种保护就是内稳态睡眠驱力。我们不知道那种保护有多少是网络过载的自然结果,有多少是大脑为防止进一步过载而做的额外努力。例如,我们可以借助咖啡因阻断内稳态睡眠驱力的腺苷组分来改善认知功能,这证明大脑提供了某种程度的网络过载保护。过载将导致清醒日中回忆和记忆巩固的进行性下降。
昼夜节律组分在睡眠的神经功能确立之前很久就演化了。然而,在适宜时机进行神经家务对机体很方便,例如,对人类狩猎采集者,夜晚是不活动的时段,这就是为什么把睡眠挂接到夜间时段在演化上有意义。其他动物可能做出了不同选择,然而,昼夜节律周期始终是神经优化最佳时机的良好提示。
简言之:神经优化不可避免:(1) 网络过载信号具有内稳态性质,而 (2) 适宜优化时间信号具有昼夜节律性质。
三过程模型(Three-process model)
Borbély 的双过程模型被扩展了代表睡眠惰性的额外过程 W,该模型的基础是自评困倦报告(Akerstedt 和 Folkard,199098)。SuperMemo 中使用的模型不包括睡眠惰性因子,因为它主要针对研究自由睡眠。
相位响应曲线(Phase response curve, PRC)
相位响应曲线(PRC)代表一个函数,它告诉我们一个振荡器的相位在响应选定刺激时移动多少,取决于这些刺激的时机。PRC 可用于研究昼夜节律以及其他生物、物理或电子系统(如心跳)。
例如,光的 PRC 可能告诉我们,在睡前 1 小时施加给定强度的绿光脉冲把昼夜节律周期向前推 10 分钟(相位延迟),而更高密度的蓝光可能把同一周期推 25 分钟。在主观夜晚,存在一个光不产生昼夜节律周期移动的死区。
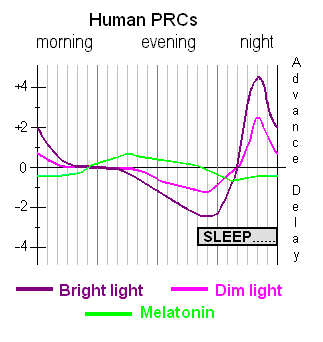
强光、弱光和褪黑素的示例人类 PRC。
不同刺激(如运动、压力激素(如皮质醇)、褪黑素等)有许多 PRC。光的 PRC 延迟侧和前移侧之间的交叉时间接近核心体温最低点。睡眠控制系统对抑制褪黑素释放的较短波长可见光最敏感(Brainard 等,2001)。
不能移动周期或影响可忽略的刺激(如一杯温牛奶)也可以绘制 PRC,然而那个 PRC 将是沿相位移动为零的水平直线。SleepChart 使用一种绘制所谓递归 PRC的算法,其中相位移动程度是相对于自由睡眠中睡眠事件实际位置测量的,不区分移动的实际原因。rPRC 因人而异,也响应生活方式变化而改变。
改变昼夜节律周期的长度(Changing the length of the circadian period)
PRC 的存在意味着时钟周期的长度处于我们的控制之下。如果我们足够早或足够晚地施加授时因子(zeitgebers),就能影响更大的相位移动,从而延长或缩短周期。每个人都可以用相对简单的措施向自己证明这一点(例如深夜强光把相位向前移,或清晨运动把相位往后移)。这就是为什么 PRC 在治疗相位移动障碍时非常重要。这也是为什么生活方式决定了相位移动和可能的睡眠问题。这就是为什么基于电力使用的现代生活方式在年轻学习一代中引发了 DSPS 的流行。最近,生物钟周期可适应性这一事实,在对宇航员能否适应火星日的研究中得到了证实(Scheer 等,200799)。此前在大鼠中也通过各种”生活方式”改变得到了证明(例如跑轮限制会增加昼夜节律时钟周期)。
相位移动的神经输入(Phase-shifting neural inputs)
我们的主时钟 SCN 受到 3 个主要授时因子输入的影响,这些输入允许相位移动:
- 视网膜下丘脑束(retinohypothalamic tract, RHT)携带光重置刺激,并通过 NMDA 受体起作用。这条输入绕过认知视觉。
- 膝状体小叶(intergeniculate leaflet, IGL)通过膝状体-下丘脑束(geniculo-hypothalamic tract, GHT)投射到 SCN,携带运动重置刺激。
- 中缝核(raphe nuclei)提供血清素能输入,据假设可调节昼夜节律周期的某些方面,并可能参与情感障碍中昼夜节律周期的变化。这条通路的损伤或血清素的减少会延长恒定黑暗中的活动相,但不影响昼夜节律周期。
递归相位响应曲线(Recursive phase response curve, rPRC)
要研究相位响应,科学家需要昂贵的实验室设备和耗时漫长的研究程序。然而,一个简单的计算技巧也使得在 SleepChart 中无需使用睡眠实验室就能观察到相位移动刺激的效果成为可能。
SleepChart 实现了**递归相位响应曲线(Recursive Phase Response Curve, rPRC)**的概念。这条曲线之所以是递归的,是因为它首先通过计算睡眠事件中的相位移动相对于用统计方法计算出的昼夜节律峰值相位(acrophase)的影响来获得。一旦获得了 rPRC 的第一次近似,它就可以用来生成主观夜晚(subjective night)中点线的更好近似,而后者又反过来用于生成 rPRC 的更好近似。经过几次这样的迭代,就足以产生与实际睡眠数据吻合良好的最佳 rPRC 拟合。SuperMemo 使用固定的 rPRC 来估计峰值相位,该固定 rPRC 大致对应于用 SleepChart 获得的 rPRC。时间生物学中使用的典型 PRC 映射的是睡眠系统对单一刺激(如光、运动、褪黑素或各种化学物质)的响应,而 rPRC 是所有自然睡眠延迟因素(包括光、活动、压力等)的综合结果。它也可以被解释为一个 PRC,其中清醒活动构成了自由睡眠系统的输入。与响应某一移动因素的 PRC 不同,rPRC 响应的是同一因素引起的晚间相位移动。因此,rPRC 并非名副其实的 PRC,任何偏离自由睡眠条件的情形都会使计算失效。rPRC 的主要优点在于,它可以从睡眠数据中推导出来,而无需采集血液样本、唾液样本或测量核心体温。这样,SuperMemo 就可以将学习与仅以普通睡眠日志数据为输入的睡眠模型相关联。
在所呈现的图中,Sleep delay (h)(睡眠延迟,小时)代表入睡时间延迟,等于实际入睡时间与 SleepChart 根据先前睡眠历史计算的入睡时间之差。由于测量参考的是自由睡眠,受自然醒来方式的影响,几乎没有相位前移的数据可用。睡眠延迟的原因可能包括光、社交互动、压力、有意识地决定延迟睡眠、运动、摄入咖啡因、药物等。
Phase shift (h)(相位移动,小时)代表相位移动,等于连续两天(发生入睡延迟的那天和次日)的两个指数加权清醒小时平均值之差。这里比较的是清醒小时而非入睡小时,因为前者较少受到实际延迟引起的内稳态移动的影响,从而更真实地反映实际的相位移动。
曲线的扁平化(与典型 PRC 相比)源于对实际睡眠数据的递归引用,这是因为 SleepChart 绘制昼夜节律峰值相位是基于同一组睡眠测量的近似。因此,多项式近似显示出相位移动随延迟增加而略有增加,这在典型的 PRC 图中并非如此。入睡时间偏离最佳入睡时间可能源于环境延迟因素,也可能源于用于绘制昼夜节律函数的启发式程序所产生的近似误差,而入睡通常自然地发生在最佳生理时间。图形固有的不对称性源于这样一个事实:提早入睡几乎总是自然的,而延迟入睡则可能是自然的也可能是被迫的。正是被迫的入睡延迟构成了自由睡眠中相位移动的主要来源。
DSPS 中的递归 PRC(Recursive PRC in DSPS)
下面呈现的图意味着,在所考虑的案例中,延迟睡眠 4 小时会导致睡眠相位移动 1.4 小时(这似乎接近最大可能的移动量)。相位前移则要求自然入睡时间比最佳就寝时间提前多达 6 小时。在最佳时间入睡会导致自然的每日延迟,在本例中为 1.0 小时,这是 DSPS 障碍或与授时因子隔离条件(如恒定照明)的典型特征。
递归 PRC 与相位前移(Recursive PRC and phase advance)
应当始终避免延迟睡眠(除非将其用作一种时间疗法)。下一张图显示了睡眠延迟如何实际上能使睡眠相位前移。这与先前描述的晚间褪黑素过量导致的相位延迟恰好相反。当清醒侵入越过昼夜节律峰值相位(即 rPRC 的稳态点之后)时,相位延迟迅速减小,直到进一步延迟睡眠会把相位往回推。自然地,这种相位操纵的”方法”对健康尤其不利,因为它意味着在主观夜晚中途清醒(参见:轮班工作和时差的健康影响)。
递归 PRC 显示的相位前移可由以下两种情况引起:(1)超过 5 小时的入睡延迟,或(2)超过 2 小时的入睡提前。
从这张示例图中,我们可以读出以下信息:
- 在自然入睡时间入睡会导致 1.3 小时的相位移动,使昼夜节律周期为 25.3 小时。
- 只有在比自然入睡时间提前 2 小时入睡时,相位移动才会被消除。
- 2 小时的相位前移是可能的,但需要不自然的提早入睡(例如由马拉松等高强度运动诱导)。
- 超过 2 小时的相位延迟不太可能发生。
- 超过 5 小时的入睡延迟可能由于清醒对主观夜晚早晨端的冲击而导致实际的相位前移。显然,这不应被视为相位延迟的”疗法”,因为这种严重的偏差会严重影响睡眠质量,并在昼夜节律周期的控制中产生重大的波动。
如果你自由运行你的睡眠,并拥有足够大的数据集(例如数月的睡眠日志),你可以用 SleepChart 中的 File : Export : Recursive PRC 生成自己的 rPRC 数据(需要 SuperMemo 15 或更高版本)。
多相睡眠中的递归 PRC(Recursive PRC in polyphasic sleep)
可以用从”Uberman 实验”中获得的数据来输入 SleepChart。显然,仅仅偏离自由睡眠条件就使得结果难以解释。即使用于获得 rPRC 的递归程序也无法有效应对主导性昼夜节律波峰的缺失。尽管如此,窥探一下”Uberman rPRC”仍然很有趣,因为它很好地反映了受多相睡眠实验影响的睡眠系统的混沌本质。
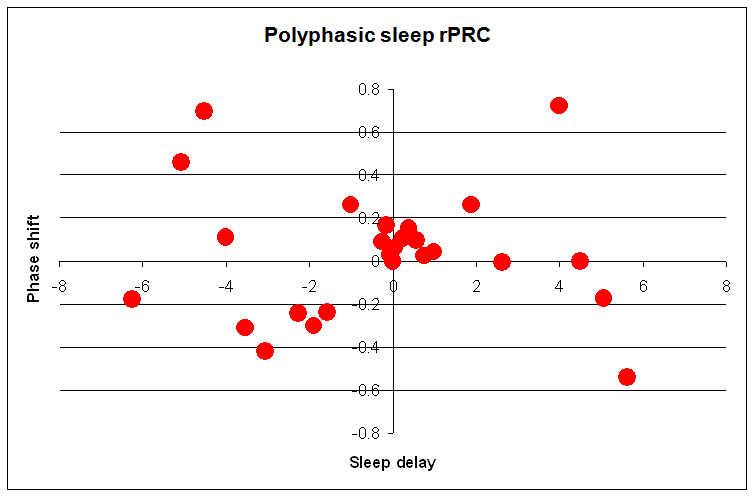
多相睡眠者基本上是随机地把睡眠相位来回推。这只能导致混乱以及所有依赖于睡眠周期昼夜节律组分的神经、内分泌和生化过程的完全失步。人们可能会预期这种混沌输入会给系统带来严重的健康后果;然而,天然的防御机制使那些试图对抗自然睡眠周期的人生活相当痛苦。结果是,那些尝试多相睡眠的人注定迟早会放弃。
混沌的相位移动输入(Chaotic phase-shifting input)
发送到相位移动输入的混沌信号(如多相睡眠中所见)可能对睡眠控制系统产生难以预测的负面后果。这一风险尚未完全知晓,也难以估计。按可能性递减的顺序,它可能包括:
- 对睡眠控制系统所发信号的脱敏
- 睡眠控制系统的长期不稳定
- 参与昼夜节律周期控制的神经细胞的损伤
第一种可能性实际上可以在轮班工作者和与睡眠剥夺持续抗争的人身上观察到。在这些个体中,“清醒头脑”和”清爽睡眠”的概念变得模糊,可以观察到他们对来自睡眠不足或在错误昼夜节律相位睡眠所导致的永久性疲劳有了更高的耐受性。换言之,某种程度的疲劳成了常态。
睡眠控制系统的不稳定也在轮班工作者中观察到。我不确定轮班引起的不稳定是否会变成慢性,还是能在相对较短的时间内完全可逆。即使在一个完美调谐的睡眠控制系统中,微小的节律扰动(如切换到夏令时)也会产生持续数天的调节涟漪。理论上,更大的扰动可能导致主振荡器与从振荡器的解耦,并需要特别缓慢的恢复才能完全稳定控制。也许这种解耦是困扰轮班工作者长期的众多疾病的主要潜在因素。
SleepChart 中的双组分睡眠模型(Two-component model of sleep in SleepChart)
SuperMemo 使用受 Alexander A. Borbély 和 Peter Achermann 的著作启发的双组分睡眠模型。与其他模型不同,SuperMemo 使用用户的睡眠数据来预测总体睡眠倾向的内稳态和昼夜节律状态。这个模型有助于在给定一天(给定特定睡眠历史)选择最佳学习时间。它也可以帮助在睡眠模式高度不规律的情况下规划最佳入睡时间。该模型不以 NREM 和 REM 睡眠事件的时机和持续时间为前提。
该模型经过调谐以拟合典型的 SleepChart 数据日志。然而,存在影响昼夜节律周期长度、内稳态警觉性下降陡度、睡眠长度偏好、睡眠结构、睡眠频谱特性、睡眠碎片化等的个体遗传差异。该模型在解释这些变量方面存在局限。如果你在模型的模拟下感到困倦,你可能更应该相信自己的直觉。如果你在模型的模拟下感到警觉,你当然可以开始学习而无视模型的预测。此外,只有当睡眠模式未被人为干扰(例如强行延迟睡眠、使用闹钟、使用药物等)时,它才是衡量你睡眠控制系统的良好指标。换言之,如果你没有自由运行你的睡眠,所呈现的模型可能无法正确映射你的昼夜节律。你可以将睡眠块标记为人为缩短或延迟(右键单击上下文菜单中的 Forced awakening 和 Delayed retirement)。然而,被标记的睡眠块效果有限,因为无法知道对睡眠模式切割的程度,因而也就无法知道人为修改睡眠所导致的睡眠控制系统扰动。
在 Borbély 模型中,睡眠的时机由代表内稳态和昼夜节律过程的曲线交叉点决定。SuperMemo 使用了一种更简单但更直观的方法,将两个组分的困倦感整合到一个总体警觉性水平中(图中的红线)。这种方法的优势在于可以从实际学习过程中获得即时反馈,其中记忆回忆水平应与模型确定的警觉性水平直接相关。将两个睡眠组分整合为总体警觉性的公式是借助在 SuperMemo 中收集的警觉性数据启发式地选择的。整合的目的是使模型预测的警觉性与 SuperMemo 中的平均回忆水平达到最佳匹配。如前所述,内稳态困倦和昼夜节律困倦都会影响 SuperMemo 中的评分,然而,只有两个组分的综合效应才能与不同内稳态和昼夜节律困倦组合下回忆的变化良好匹配。在 SuperMemo 使用的模型中,当总体警觉性降至某一水平以下时,睡眠被启动。因此,睡眠可以由两个困倦组分独立启动,但对于内稳态和昼夜节律睡眠倾向变化的不同组合,睡眠的时机和持续时间会有所不同。尽管使用了不同的方法来确定睡眠启动,但在所研究的案例中,模型的预测与自由睡眠条件下的实际睡眠日志数据相当吻合。
要查看模型对你自己睡眠数据在某一天的预测,请确保你在 SleepChart 中填写了最近几天的睡眠日志,并在睡眠日志中按住 shift 单击有问题的那一天。
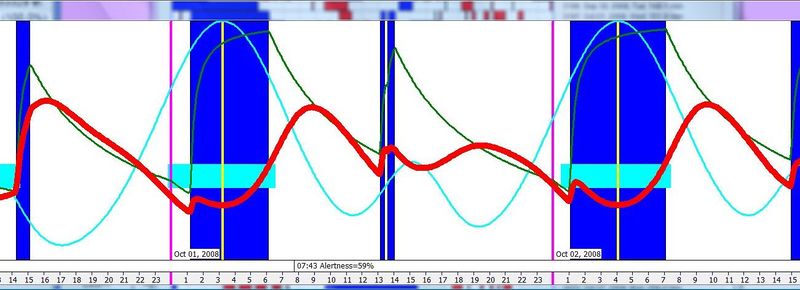
SuperMemo 中的双组分睡眠模型:横轴代表时间。蓝色块显示实际睡眠事件。水绿色线显示带有中午午睡(siesta)小峰的 24 小时昼夜节律睡眠驱动力。绿色线是内稳态睡眠驱动力的倒数,可以解释为内稳态警觉性,它在清醒期间指数下降,并由慢波睡眠快速恢复(为简化起见,如 Borbely 模型中一样,假设整个睡眠块的贡献与其长度成正比,因为 SleepChart 模型不考虑睡眠阶段)。黄色竖线显示昼夜节律峰值相位(昼夜节律的夜间中点峰值)的预测。峰值相位计算借助相位响应曲线模型完成(而非早期 SleepChart 版本中使用的统计模型)。红线显示合成的警觉性(峰值最适合学习,谷值最适合睡眠)。例如,2008 年 10 月 1 日上午 7:43 的警觉性被预测为最大值的 59%,但在清醒后的头 2 小时内会快速上升(这是夜间睡眠被过早终止的典型症状)。图片显示 2008 年 10 月 1 日有两个警觉性峰值,分别在上午 9 点和晚上 7 点。这些时段可能最适合当天的学习。
要在 SuperMemo 中查看你自己更准确的内稳态和昼夜节律警觉性呈现,请参见 SleepChart 中的 Alertness 标签页。
REM 反弹假说(REM rebound hypothesis)
研究者们知道,Borbély 的双过程模型并不完整,无法解释所有已知的睡眠特性,甚至无法解释所有可能的睡眠模式(例如各种打盹习惯、新生儿睡眠、不规律睡眠模式、精神障碍中的睡眠等)。人们曾多次尝试通过在特定情况下可能显现的新变量来扩展该模型(例如添加噪声来模拟自闭症儿童的睡眠-清醒模式,用超日节律动力学来建模 NREM-REM 的发生,添加光照强度的影响等)。Borbély 和 Achermann 一直在研究有助于使模型更完整的各个方面。他们的研究目标之一是 REM 睡眠剥夺之后的 REM 睡眠反弹。人们早就发现,REM 睡眠剥夺会减少 α 活动、清醒和 NREM 睡眠。这些是 REM 内稳态代偿的清晰迹象(Borbely 等,1990100;Brunner 等,1993101)。已经有人提出,将 NREM 中肌张力丧失事件(MAN)的增加作为 REM 睡眠压力增加的标志(Achermann 等,2002102)。
异常打盹(Kinky napping)
多年来,我在 SleepChart 日志中观察到一个用双过程模型无法解释的异常现象。在睡眠不规律的人中,晚间的小睡通常异常漫长且不能恢复精力。这些长小睡清除了睡眠倾向的内稳态组分,并经常导致更晚的入睡时间。在某些极端情况下,这可能导致对最佳睡眠时机的混淆。受影响者会小睡足够长时间,以至于失去对自己主观夜晚时机的感知。下图展示了这样一个经典案例。
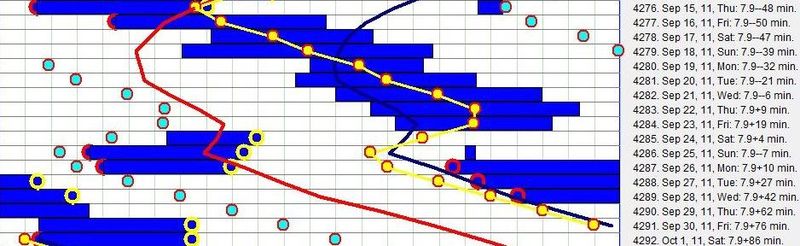
“异常”打盹可能导致 DSPS 中对主观夜晚时机的混淆。
在上面的示例睡眠日志中,一位在家工作的中年女性患有延迟睡眠相位综合征,显示出从凌晨 2 点入睡到睡过正午的清晰且相当规律的相位推进。这位女士声称自己睡眠不规律、白天疲倦,且永远不知道何时入睡才能获得”一夜好眠”。9 月 24 日,由于感到疲倦,她在上午 6:30 去小睡。这次小睡出乎意料地持续了 3.5 小时,并产生了当天不需要更多睡眠的印象。尽管有些疲倦,这位女士当晚并没有上床睡觉,尽管图表清楚地说那是她的主观夜晚期间,她应该就寝。经过一个特别疲惫的傍晚和夜晚后,这位女士在凌晨 3:30 入睡,以为这是她的”夜间睡眠”。那次睡眠持续了 6 小时,足够清爽以”冒充”夜间睡眠。这完成了夜间与午睡时段之间的角色互换。两个昼夜节律低谷在睡眠者的心中被互换了。这种互换随后反映在就寝仪式、期望和其他习惯中,尽管睡眠可能极度不清爽,这些习惯仍可使角色反转持续数天。然而,夜间与午睡的反转并不稳定。9 月 25 日,在主观夜晚期间,又进行了一次短小睡,似乎特别清爽。但这种清爽印象很快消散,第三个”在午睡时间的夜晚”随之而来。9 月 26 日,足够的睡眠债务累积导致一次”小睡”突然变成了 8 小时深沉清爽的睡眠。经过 3 次延长的小睡事件后,睡眠模式翻转回常态。这种角色互换无法用双过程睡眠模型解释,也无法用 SleepChart 中使用的模型解释。这三次在午睡时间之后进行的异常小睡,由于没有得到昼夜节律午睡低谷的增强,本应更短。这些小睡也发生得太早,无法利用夜间昼夜节律低谷。换言之,这些”异常”小睡尽管缺失了睡眠的昼夜节律组分,却持续了异常长的时间。
REM 亏欠(REM deficit)
在睡眠模式中数十次看到这些”扭结”后,我开始相信,睡眠倾向的 2 过程模型需要通过第三个组分来扩展。然而,很难想出一个能合理契合我们关于睡眠功能及其演化的已知内容的合理假说。一个重要线索来自受异常小睡影响者的访谈。这些长小睡似乎经常由饮酒触发,在某些情况下则是吸食大麻。如果酒精或大麻的施用时机与主观夜晚前不久的晚间清醒时段一致,第二天就可能出现异常小睡。此外,这些小睡之前都有一个特别强烈的早晨不清爽感,这在酒精或大麻滥用中很常见(使用安眠药甚至褪黑素时也是如此)。由于这两种物质都已知会减少 REM 睡眠占比,我假设正是 REM 睡眠的亏欠导致了上述睡眠扰动。
我还记录了一些案例,异常小睡发生在不涉及酒精、大麻或其他影响睡眠物质的、健康且清爽的夜间睡眠之后。这些其余案例也有另一个共同因素:最佳入睡时间的一次性大幅延迟及由此产生的睡眠相位移动。这与 REM 亏欠假说一致。如果睡眠延迟超过昼夜节律 REM 峰值,其 REM 占比也已知会降低。
最后,这些异常小睡与健康且时机恰当的小睡不同,被报告为梦境丰富。这也可能表明它们可能涉及 REM 代偿功能。
如果 REM 亏欠假说成立,我们就需要始终分别考虑内稳态 REM 和 NREM 睡眠倾向。在健康睡眠中,REM 组分可能难以察觉。一些研究者假设内稳态 REM 驱动取决于先前的 NREM 睡眠。如果是这样,从内稳态角度看,健康的夜间不会产生 REM 亏欠,而清醒活动只会产生内稳态 NREM 困倦。
REM 亏欠如何产生这些延长的小睡?有一些迹象表明 REM 睡眠也能增加对 NREM 睡眠的需求。因此,这两个功能至关重要的睡眠阶段可能产生一个相互放大的循环,直到两个睡眠组分的需求都得到满足才会停止。为什么 REM 睡眠会在正常夜间睡眠的末期增加?其中一些增加是昼夜节律性的,另一些可能来自内稳态 NREM 睡眠需求更快得到满足这一事实。对以 REM 终止的睡眠的生物学解释是困难的,尤其是在 Buzsáki 关于 REM 中海马体”训练”模型的背景下。带着一张白纸醒来似乎在生物学上更有利。也许最后一个 REM 时段负责了清晨的创造性突破?只有详细的数学建模和与实际睡眠周期测量的比较,才能回答关于 NREM 和 REM 睡眠之间内稳态相互作用的问题。
睡眠倾向的三个组分(Three components of sleep propensity)
SleepChart 无法轻易验证 REM 亏欠假说的本质。不仅其日志中缺少睡眠阶段,在 SuperMemo 或 SleepChart 用户中,在家庭条件下检测 REM 睡眠也不切实际。然而,解释睡眠日志中异常小睡所需的第三个变量——我称之为 RD 变量(代表 REM 亏欠)——可能被纳入双过程模型,以期能从数学上解释”扭结”对估计睡眠相位的影响。如前所述,这些异常小睡并不需要酒精或其他抑制 REM 的因素,被标记为 Delayed retirement 的睡眠块由于相对于昼夜节律 REM 峰值的错误相位对齐,常常也会产生类似效果。一旦睡眠失步能用 RD 变量成功解释,就该由睡眠实验室使用 EEG 测量来验证模型了。RD 变量与其他两个睡眠变量(H 和 C)之间的相互作用并不简单。例如,高 RD 不足以启动睡眠,因为没有 H 和 C 的适当组合就不可能启动睡眠。高 RD 和高 C 也可能不足(正如在异常小睡之后的夜间睡眠相当短这一事实所表明的,原因是低 H)。然而,高 RD 和高 H 可以启动充分的睡眠,并导致异常小睡,对随后的夜间睡眠(低 H)和睡眠相位产生可能的负面后果。在撰写本文时,我仍不确定睡眠相位是如何受到影响的,但我相当确信确实如此。例如,在前面给出的例子中,睡眠相位似乎被往后移了几个小时,然而,这也可能是由 SleepChart 中所用模型的缺陷所致(恰恰是由于缺失了 RD 变量)。低 C 时需要高 H 和高 RD 才能启动睡眠这一点,与当前关于 NREM 和 REM 睡眠阶段相互作用的最新研究一致,即一个阶段增加对另一个阶段的需求。
睡眠-清醒双稳态开关(Sleep-wake flip-flop)
睡眠-清醒双稳态开关是一个由两组脑核团组成的系统,它产生从睡眠到清醒的快速切换,反之亦然。一组核团负责诱导睡眠并抑制唤醒中心,而另一组则起相反作用。两组相互抑制。这意味着到了该睡觉的时候,睡眠中心占据上风并关闭清醒中心。之后,在早晨,清醒中心接管控制并关闭睡眠中心。这个睡眠-清醒开关的构造方式使得从睡眠到清醒的过渡以及反向过渡都相当快速而彻底。在健康的睡眠周期中,我们在睡前只有很短的时间处于半清醒状态,而在早晨也许稍长一点。不幸的是,这并不意味着我们可以随心所欲地切换这个开关。它也不意味着我们在睡前不会感到疲倦。内稳态困倦的增加是一个自然过程,它贯穿整个清醒时段。只有从清醒到睡眠的过渡才是快速的,而且是在内稳态困倦遇到足够程度的昼夜节律困倦之时。睡眠-清醒双稳态开关由食欲素(orexin)神经元稳定。正如 Siegel 所证明的,食欲素(也称下丘脑分泌素 hypocretin)的水平与昼夜节律周期无关,而与特定行为有关。在清醒活动(例如运动)期间,食欲素水平可能保持较高,从而阻止睡眠-清醒开关的切换。当食欲素稳定器失效时,发作性睡病就会出现,开关变得不稳定,导致在难以预测的情况下一天内出现多次睡眠事件。
睡眠-清醒双稳态开关最重要的组成部分是:
- 睡眠侧的腹外侧视前核(VLPO),以及
- 唤醒侧的结节乳头核(TMN)、蓝斑(LC)和中缝背核(DR)。
一旦睡眠启动,另一个开关开始运作:负责 NREM 和 REM 睡眠之间转换的那个开关。
视交叉上核(SCN)
人脑中 harbors 一个以略长于 24 小时的周期运行的时钟。这个时钟被称为视交叉上核(SCN),位于前腹侧下丘脑。SCN 由两组神经元(各约 10,000 个,0.25 mm³)组成,双侧对称地位于视交叉正上方。SCN 在女性中略长,两性之间在 VIP 表达神经元上存在显著差异(男性最多可达两倍)(Swaab 等,199059)。同性恋男性的 SCN 更大,VP 表达神经元数量是异性恋男性的两倍(Swaab 和 Hofman,1990103)。顺便说一句,我相当确信这种差异并非出于选择,也无法通过自律或祈祷来矫正。
1972 年,SCN 被确定为人体可以在没有环境线索的情况下运行并从视网膜接收重置输入的主时钟。SCN 中的生物钟基因负责一个基因表达的昼夜节律周期,决定了 SCN 的输出。SCN 中的神经元表达这一周期,并以各种神经和激素形式反映在从 SCN 传递到其他脑核团和身体其余部分的信号中。如果我们手术损伤 SCN,昼夜节律周期会减弱或消失。它可以通过 SCN 细胞移植来恢复。
SCN 信号在主观白天最活跃,尤其是在傍晚时段。它在主观夜晚最弱,尤其是在体温达到最低值的清晨。如果你曾尝试过多相睡眠,正是 SCN 会困扰你,让你渴望核心睡眠,并让你在主观夜晚时段过度睡眠。SCN 控制警觉性、注意力、激素释放、体温、褪黑素分泌、进食等。SCN 的大部分输出流向室旁下区(SPZ)和下丘脑背内侧核(DMH)。背侧 SPZ(dSPZ)中的神经元影响体温的昼夜节律,而腹侧 SPZ(vSPZ)中的神经元运行清醒和睡眠周期。vSPZ 进而向 DMH 发出指令,后者是清醒行为、运动活动、皮质醇周期、进食等的首席指挥中心。DMH 影响促眠的 VLPO 和促醒的 LHA(下丘脑外侧区)。对 VLPO 和 LHA 的损伤可分别导致失眠或无法抗拒的嗜睡。
SCN 和 DMH 在控制行为的最基本、最有影响力的神经通路十字路口的这种中心位置,有力地展示了少数几千个神经元如何对作为活跃进食和求生的我们所做的事施加强大影响。这应提醒每个人,睡眠卫生对这个人脑中微小结构的正常运作至关重要。用闹钟、轮班工作等扰乱昼夜节律周期可能导致一连串身心疾病。关于 SCN、DMH 和身体其余部分之间相互作用的详尽综述,参见 Clifford B. Saper 博士的”Hypothalamic regulation of sleep and circadian rhythms”(Saper 等,2005104)。有趣的是,Saper 博士假设正是 DMH 整合了运动或社交互动等昼夜节律重置刺激。在啮齿动物中,DMH 还可被食物可得性甚至温度重置。不过,你不太可能借助食物或温度等授时因子来对抗时差或适应任何轮班模式。
SCN 以略长于 24 小时的周期振荡。为适应 24 小时的世界,振荡需要每日重置以匹配地球的日光周期。重置借助授时因子(“时间给予者”)完成,如光、运动、进食等。最重要的授时因子是光。光信号由视网膜中表达黑视蛋白的谷氨酸能视网膜神经节细胞(pRGCs)接收,并从那里通过视网膜下丘脑束(RHT)传递到 SCN。光信号和其他授时因子对昼夜节律相位的影响由所谓的相位响应曲线(PRC)描述。最重要的是,晨光信号有助于重置周期,昼夜节律周期缩短以匹配 24 小时日光周期。借助授时因子,周期略长的振荡器被 SCN 介导的微小重置拉回与日光同步,从而提供稳定的振荡。无法有效引导其振荡器的人患有相位移动障碍。患有 DSPS 的人可以尝试使用调光器、在晚上放慢节奏、恰当安排的运动和早晨的强光。患有 ASPS 的人应采取相反的措施(例如晚间 3000 勒克斯的光照)。除光之外,SCN 还受活动影响。运动活动通过激活膝状体小叶(IFL)中含 NPY 的神经元和中缝正中核(MRN)中的血清素能神经元来影响 SCN。这就是为什么运动和社交互动能作为强大的授时因子。
当我们使用人工照明或进行刺激性的晚间活动(如看电视、上网、玩电脑游戏、阅读等)时,SCN 指挥的神经交响乐就会出乱子。有人假设,在主观夜晚,光(以及其他刺激)可能以两种不同方式影响 SCN。短光脉冲简单地改变生物钟基因的表达,并导致沿 PRC 的相位移动。然而,持续照明可能导致 SCN 神经元与受 SCN 影响的下游核团之间的解耦,从而产生危险的节律紊乱(Ohta 等,2005105)。在轮班工作或时差中看到的对昼夜节律周期的持续干扰,可能像啮齿动物研究所示那样导致渐进的精神衰退(Ree 等,1985106)。与衰老和阿尔茨海默病相关的昼夜节律变化可能与 SCN 中细胞的丢失或其输入的变化相关。表达血管加压素的细胞在阿尔茨海默病中的减少尤为突出。所有形式的睡眠周期人为控制,包括使用闹钟,都可能影响这些珍贵神经元的健康。
SCN 向背侧 PVH(细细胞室旁核)发送投射,后者的神经元投射到脊髓中的交感节前神经元,进而影响松果体和褪黑素的释放。这告诉我们,经常被宣传为”天然安眠药”的褪黑素,是由在 SCN 控制下游的松果体产生的。这就是为什么它不能被视为控制昼夜节律困倦的核心因素。褪黑素确实沿其独特的褪黑素 PRC 产生相位移动(参见 PRC 部分的图)。这种效应可能由褪黑素对 SCN 的直接影响引起。然而,提早入睡也会导致更早醒来,这同样会产生相位移动效应。
背内侧下丘脑核(DMH)
Saper 博士及其同事证明,对大鼠下丘脑背内侧核(DMH)的兴奋毒性损伤会导致昼夜节律的严重障碍(Saper 等,2003107)。由于损伤的动物睡得更多,有人提出 DMH 的影响主要是激活性的,尽管对现有发现的其他解释也是可能的。看来,SCN 的大量输出经由室旁下区(SPZ)到达 DMH,然后才通过抑制性通路到达负责启动睡眠的 VLPO。DMH 还投射到含有促醒食欲素神经元的下丘脑外侧区(LHA)。有人假设 DMH 可能处于控制沿昼夜节律周期变化的各种变量(如觉醒、进食、运动活动、皮质醇水平、体温、褪黑素等)的中心。限制性喂食使 DMH 的昼夜节律同步,使最高的 c-Fos 表达和运动活动与进餐时间重合(Saper 等,2006108)。由于到达 DMH 的大部分输入经由 SPZ,重要的是要注意 SPZ 的背侧和腹侧部分似乎发挥不同功能。dSPZ 的损伤减少体温的昼夜节律,而 vSPZ 似乎控制睡眠-清醒周期和运动活动(Saper 等,2001109)。dSPZ 通过内侧视前区(MPO,包括正中视前核和腹内侧视前核)控制体温。DMH 受控制食欲的激素饥饿素和瘦素经腹内侧核(VMH)和弓状核(ARC)的影响。VMH 增强脂肪组织中的脂解并减少进食。Saper 博士假设 DMH 可能作为次要的昼夜节律控制中心,使节律能够驯化到食物的可得性。然而,从控制系统的角度来看,相位移动 SCN 而非采用第二个异步或相位锁定的振荡器在生物学上似乎更合理。在人类中,很难通过除使用各种授时因子(其中食物是非常弱的一个)进行微小相位移动以外的方式影响昼夜节律周期。因此,轮班工作者或时差旅客似乎不太可能从改变饮食时机中获得实质性益处。大鼠和人类之间的差异自然不能排除。尽管如此,DMH 无疑是一个非常有趣的进一步研究目标。
腹外侧视前核(VLPO)
腹外侧视前核(VLPO)是启动睡眠和维持慢波睡眠所需的主要大脑中心之一。该区域的损伤会使睡眠减半,并导致失眠伴持续疲倦。根据损伤类型的不同,NREM 和 REM 都可能受到影响。由于这一作用,VLPO 常被称为”睡眠开关”。在夜行性和昼行性动物中,SCN 在白天活跃,而 VLPO 主要在睡眠期间活跃。一旦 VLPO 开启,据信它会维持对保持大脑皮层”清醒”的单胺能和胆碱能兴奋系统的抑制。这些 VLPO 投射到达结节乳头核(TMN)(组胺)、下丘脑外侧区-穹隆周区(LHA/PF)(食欲素)、腹侧导水管周围灰质(vPAG,多巴胺)、蓝斑(LC)(去甲肾上腺素)、臂旁核(PBN)和中缝背核(DR)(5-羟色胺)、外侧被盖(LDT,乙酰胆碱)和脚桥被盖核(PPT)(乙酰胆碱)。VLPO 实施的抑制由 GABA 能神经元以及对组胺能结节乳头核(TMN)的甘丙肽能输入介导。对 TMN 和其他警觉核团的抑制导致警觉激素下降和皮层激活下降,引起困倦。一个假说说,VLPO 的不同群体可能负责表达 NREM 和 REM 睡眠的昼夜节律方面。VLPO 细胞的一个子集能够刺激 LDT 和 PPT 中的胆碱能神经元。这有助于诱导 REM 爆发,在 REM 睡眠期间激活皮层而不产生清醒。
VLPO 通过下丘脑背内侧核(DMH)从 SCN(主生物钟)接收其昼夜节律信号输入,DMH 是另一个通常与 SCN 同步的大脑时钟。VLPO 神经元不会建立睡眠的内稳态需求,然而,一些内稳态机制(如腺苷的胞内积累)可能抑制胺能或胆碱能唤醒中心,从而激活 VLPO。例如,向基底前脑输注腺苷激动剂可同时增加 NREM 和 REM 睡眠(Satoh 等,1999110)并增加 VLPO 中的 c-Fos(Scammell 等,200193)。在睡眠剥夺中,VLPO 的活动不比普通清醒时高多少。这种低活动水平一直持续到就寝时间。一旦睡眠开始,在睡眠剥夺条件下 VLPO 神经元放电率可能翻倍(Saper 等,2005104)。这表明,即使 VLPO 不建立内稳态睡眠倾向,它最终还是受内稳态机制的影响。这也表明 VLPO 位于昼夜节律和内稳态信号整合器的下游。胺能唤醒核团如 TMN、LC 和中缝核构成睡眠-清醒开关的一部分。该开关由来自下丘脑外侧区-穹隆周区(LHA/PF)的食欲素/下丘脑分泌素细胞稳定,尤其是在运动活动或进食期间。因此,尽管存在竞争性抑制影响,唤醒仍可不受中断地维持。唤醒核团在清醒时抑制 VLPO,正如 VLPO 在睡眠时反过来抑制它们。科学家们认为,这种相互抑制构成了一个具有尖锐状态转换的经典非平衡双稳态开关。这正是帮助我们快速入睡、快速醒来、在过渡中花费最少时间、在期望状态(警觉或深睡)中花费最多时间的原因。
孤束核(NTS)
这一节更多地讲述科学中的潮流和时尚,而非孤束核(NTS)在睡眠中的实际参与。我在 20 世纪 80 年代初大学学习生物学期间听说了 NTS 对睡眠的重要性。一段时间后,我在 SuperMemo 中添加了几条关于 NTS 的条目,以巩固这些知识。2000 年撰写Good sleep, good learning, good life一文时,我仍然提到了 NTS 以及摇晃婴儿入睡可能如何起作用,尽管我知道 NTS 的破坏不会导致失眠——这本应是一个重大线索。NTS 似乎更多地参与处理来自肠道的信号,而这些信号在睡眠控制中只起次要作用。最终,我成了自科学诞生以来就困扰科学的同一种痼疾的受害者。有时需要老一代科学家凋零,新观念才能扎根。旧知识使我们更加保守,因为不知道会驱使我们寻找答案,而知道——即使我们的答案是错的——会使我们变得被动。一旦你相信自己知道所有答案,调查的压力就会减少。归根结底,许多其他大脑中心扮演着与 NTS 类似的角色。毕竟,大脑是一个高度连接的结构,中枢神经系统某一角发生的事情很少对其他角落的事件没有影响。因此,几乎所有主要核团的激活都会在网状激活系统内产生唤醒或抑制效应——而该系统多年来一直是我们思考唤醒问题的主要支柱。与此同时,回到 2000 年,我几乎没有提到 VLPO,因为它可能不够时髦。我们可能会也可能不会面临与 SLD 类似的情况,SLD 已作为公认的 PPN/LDT REM 开启系统的替代方案出现。即使是 DMH 也可能无法免受时尚的影响。像 NTS 一样,它也参与进食行为。也许十年后,这篇文章将需要彻底重写,许多旧时尚会消逝。同样,在所有新科学发现触手可及、我们可以通过电子邮件和其他方式轻松交流的时代,我们都会表现出较小的随大流倾向。更重要的是,新的研究技术可能会开辟仍可能受时尚影响的新领域,而本文讨论的主题将逐渐凝固和固化。
顺便说一句,时尚的痛苦曾经是推动我远离同行评审写作、转向博客的主要因素。撰写本文是一种极致的乐趣。本文使用增量写作写成,并作为 wiki 协作打磨。作为商业公司的一员,我不受”不出版即灭亡”压力的束缚,这是一种宝贵的自由。早在 1992 年,我和 Gorzelanczyk 博士研究了间隔效应的文献,并得出结论:我们用 SuperMemo 收集的数据,以及对间隔重复概念的清晰计算表述,将席卷教育和记忆科学界。当时绝大多数间隔效应文献都聚焦于短期研究(例如只在试验后一周检查记忆效应)。Bahrick 对西班牙语词汇保持的研究是一个重大的杰出例外。然而,Bahrick 只能在原始训练多年后研究词汇的保持,在学习期间或测量前的漫长时期内,没有关于单个词汇暴露时机的具体数据。在此背景下,我们认为我们拥有启动学习和记忆科学新革命所需的一切。令我们极度失望的是,我们无法将论文推过 Memory and Cognition 的发表关。我们的失败部分源于经验不足和缺乏资历——我们俩都刚刚从大学获得硕士学位。无论我们多么努力地围绕时髦的间隔效应措辞论文,我们都无法将其塑造成符合记忆科学主流的形态。我们从一本不知名的旧期刊中为我们的重复调度方法挑选了听起来最科学的名字——我们称之为 repetition spacing。这个术语后来演变为间隔重复(spaced repetition),并仍是那篇原始论文唯一有形的遗产,尽管它尚未渗透到学术命名空间。也许永远不会。更通用的”distributed spacing”或”distributed presentation”仍然占主导地位,而计算机化抽卡的用户现在一致地说”spaced repetition”。Memory & Cognition 的编辑祝贺我们的结果,并提到论文的计算方面使其适合专注于计算机算法的期刊。记忆科学的时尚世界与我们的主张如此不同,以至于没有顶级模特接受我们的立场。最终,我们在较不知名的 Acta Neurobiologiae Experimentalis 上发表了。可悲的是,这篇论文在发表 20 年里只获得了 18 次引用,而当它被提及时,也是谨慎引用的。毕竟,“优化算法”感觉像一个黑箱。它免费提供给任何人,但很难在运行中研究它。它不是一个整洁的公式,而是一个算法,最好在计算机上运行并用计算机手段研究。我们的这一款服饰显得极不时髦。如今它已被数百万人普遍使用,每月都有新设计师加入。不过,科学界目前仍基本不受影响。它在等待一批在间隔重复滋养下成长的新人才。
科学中的时尚是我们集体认知偏见的一部分,它们拖慢进步,不可避免,但从长远来看,它们会回归到近似真理的均值。重要的是,每后退一步,我们都能前进十几步。
腺苷(Adenosine)
腺苷是内稳态睡眠驱动的内源性标志物之一。在清醒期间,随着皮层和大脑其他部分不断燃烧其糖原储备,ATP 被转化为腺苷并在细胞外积累。腺苷的作用最初是在发现其全身给药可促进睡眠后被揭示的(Radulovacki 等,1984111)。
基底前脑胆碱能神经元活动的增加导致腺苷的积累,腺苷反过来通过其 A1 受体抑制该区域的活动(Strecker 等,2000112)。这是睡眠的内稳态触发因素之一。腺苷在基底前脑的积累尤为重要,因为其效应在这里最为显著(Strecker 等,2000112)。基底前脑中的积累导致某些胺能唤醒中心的抑制和 VLPO 的去抑制,从而促进睡眠。向喙侧基底前脑输注 A2A 受体激动剂可同时增加 NREM 和 REM(Satoh 等,1999110)。此外,腺苷 A2A 受体激动剂刺激 VLPO。由此产生的 VLPO 激活可通过 c-Fos 活性的增加来测量(Scammell 等,200193)。上述某些情景最近受到了质疑,因为发现基底前脑胆碱能神经元损失 95% 的大鼠尽管缺乏标志性的腺苷增加,却表现出完好的睡眠稳态。
腺苷特别有趣,因为其著名的拮抗剂是咖啡因。咖啡因与腺苷受体结合,从而阻断内稳态睡眠倾向。这证明网络过载不是内稳态睡眠驱动的原因,或至少不是唯一原因。大脑已经进化出睡眠保护机制(此处涉及腺苷),以确保在网络过载导致任何显著后果之前,内稳态睡眠驱动推动动物进行一次睡眠休息并完成必要的神经内务。
NREM 和 REM 睡眠(NREM and REM sleep)
NREM 和 REM 的交替(NREM and REM alternations)
在夜间,我们交替进入两个睡眠阶段:
使用 EEG 测量,科学家能够区分对应于逐渐加深的 4 个 NREM 睡眠阶段。在较新的文献中,你可能会读到三个阶段,因为 NREM 的第 3 和第 4 阶段被合并为单一的慢波睡眠阶段。

一夜中典型的 NREM-REM 循环。
当我们闭上眼睛,需要 3-15 分钟进入第 1 阶段 NREM 睡眠(在健康且调节良好的个体中)。在这个阶段我们经常会体验到与坠落感相关的小抽搐。轻微的干扰就会把我们唤醒,我们甚至会否认自己睡着了!一旦第 1 阶段 NREM 巩固,我们就会进入仍然相对较浅的第 2 阶段 NREM 睡眠。之后我们进入第 3 阶段(和第 4 阶段)NREM,也称为深睡眠或慢波睡眠(SWS)。
历史上,REM 睡眠对记忆和学习的重要性在我们真正意识到慢波睡眠的作用之前就已得到记录。因此,关于睡眠的文章和书籍中充斥着对 REM 睡眠在学习中作用的过度强调(与 SWS 相比)。随着时间的推移,REM 剥夺研究受到了大量批评。今天我们知道,不间断 NREM 和 REM 睡眠的自然和谐交互对记忆、学习和创造力至关重要(Salzarulo 等,2000113。
残酷的睡眠剥夺研究实际上表明,被剥夺睡眠的大鼠如果被剥夺 REM 比被剥夺 NREM 活得更久。被剥夺睡眠的大鼠存活 2-3 周。仅被剥夺 REM 睡眠的大鼠存活约五个月。
打盹的人类受试者报告说,第 4 阶段 NREM 感觉最具恢复力。去甲肾上腺素、5-羟色胺和组胺的释放在 REM 期间被抑制。在做梦期间,初级视觉皮层不活跃,而其次级区域活跃。这类似于人们被要求想象一个视觉场景而非实际看到该场景的情况。盲人的梦境更偏听觉和触觉。这似乎证实了 REM 睡眠在经验回放和记忆优化中的作用。他们在 REM 睡眠中也不表现出典型的眼动模式。这些观察导致了一种观点,即 REM 睡眠对创造力至关重要(比 NREM 睡眠更重要)。在 REM 期间,胆碱能调节抑制了从海马体到新皮层的信息流。这被认为有助于在新皮层区域内建立新的关联。
NREM 和 REM 的演化(Evolution of NREM and REM)
REM 睡眠在系统发生上比 NREM 睡眠年轻。鱼类、两栖动物或爬行动物不表现出典型的 REM 睡眠。然而有趣的是,REM 睡眠同时存在于哺乳动物和鸟类中。这使一些科学家假设 REM 睡眠被演化发明了两次!显然,REM 睡眠对具有鸟-哺乳动物智商水平的生物的生存起着关键作用(参见:动物睡多少?)。然而,研究过针鼹中 REM 睡眠的 Siegel 博士得出结论,这种动物的睡眠结合了 REM 和 NREM 睡眠的方面。因此,他提出 REM 和 NREM 可能是从系统发生上更古老的统一睡眠形式演化而来(Siegel 等,1996114)。如果 REM 睡眠在其功能上与 NREM 截然不同,并且如睡眠中的神经优化相关理论所暗示的那样复杂,那么这种”再发明”因素可能被演化论者用作反对神经优化的论据。然而,像空中飞行一样,再发明加上复杂性同样可能增加对睡眠关键神经功能重要性的份量。
NREM 和 REM 的亏欠(NREM and REM deficits)
残酷的睡眠剥夺研究实际上表明,被剥夺睡眠的大鼠如果被剥夺 REM 比被剥夺 NREM 活得更久。被剥夺睡眠的大鼠存活 2-3 周。仅被剥夺 REM 睡眠的大鼠存活约五个月。睡眠剥夺之后,首先偿还的是 SWS 亏欠。SWS 亏欠是 NREM 睡眠剥夺的结果。REM 亏欠随后偿还。一个常见的情景是 SWS 亏欠在恢复的第一晚完全偿还,而 REM 睡眠亏欠可能持续到第二个恢复夜晚。
强制失步协议(forced desynchrony protocols)是将受试者的睡眠与其自然昼夜节律周期解离的睡眠协议。在强制同步条件下,我们可以观察到,表征 NREM 睡眠的慢波活动与内稳态睡眠倾向相关,而睡眠事件中 REM 睡眠的比例同时取决于内稳态压力和昼夜节律周期。
NREM 控制(NREM control)
NREM 睡眠主要通过内稳态机制控制。在清醒的一天中,我们建立起启动睡眠及其更深的 NREM 阶段的压力。如果在没有昼夜节律组分参与的情况下启动睡眠,它很可能短而仅含 NREM。与内稳态困倦相关的信号之一是腺苷的积累。正是腺苷受体受到咖啡因的影响,从而产生其对降低内稳态睡眠压力的短暂效果。腺苷积累的后果之一是胺能唤醒中心的抑制、基底前脑的抑制,以及负责启动睡眠的首席大脑中心 VLPO 的去抑制。REM 睡眠也有内稳态组分,然而在亏欠时期,首先补偿的是 NREM 睡眠亏欠。还有更多证据表明 REM 睡眠增加 NREM 睡眠压力(Beersma 等,1990115)。除腺苷外,其他信号如白细胞介素-1、肿瘤坏死因子、干扰素、前列腺素 D2、NO、GHRH 等也与内稳态睡眠倾向的增加相关(Krueger 等,2008116)。
睡眠剥夺同时增加 NREM 和 REM 睡眠倾向。短睡眠者的 NREM 2 较少,但关于他们睡眠的实际质量和有效性的数据很少。Thomas Edison 或 Nikola Tesla 以睡得相对较少而闻名,而 Einstein 是著名的长睡眠者,据说每晚睡超过 9 小时。有趣的是,所有这些天才也都属于著名的打盹者。少睡确实能压缩不太关键的 NREM 2 睡眠,但没有证据表明这能成为你的常规习惯而不损害 NREM-REM 交互的质量。根据目前可用的睡眠数据,结论是:不要试图通过少睡来压缩 NREM 2。你很可能会损害睡眠中发生的记忆优化过程。
一些科学家认为,在睡眠期间,中脑-脑桥交界处的一个超日节律振荡器控制着 NREM 和 REM 睡眠的规律交替。然而,“振荡器”一词相当误导,因为 NREM-REM 相互作用的机制更像是一种双稳态开关性质,且交替的时机相当不规律,表明存在最终导致睡眠周期消亡的显著内源性和外源性内稳态影响。
神经调节(Neuromodulation in sleep)
睡眠的 AIM 模型(Hobson 等,200089)强调了在胺能活动缺失时主导 REM 的胆碱能调节的重要性。
在 NREM 睡眠中,脑干和前脑中的胆碱能系统不如清醒时活跃。血清素能中缝核和去甲肾上腺素能 LC 也较不活跃。另一方面,在 REM 睡眠中,这些胺能结构被强烈抑制,而胆碱能系统活跃起来。组胺的释放在 REM 中也下降。有人假设,胆碱能调节抑制了从海马体到新皮层的信息流。这被认为在双网络学习模型中扮演重要角色,在该模型中,海马体基于旧信息建立新的关联(参见:睡眠中的神经优化)。
不相信睡眠在记忆和学习中作用的 Siegel 博士认为,REM 睡眠服务于血清素能、去甲肾上腺素能和组胺能神经元停止放电后的恢复。仿佛它们被过度使用,试图补充其资源。这种解释可能通过关闭检验(参见:睡眠理论),因为许多这类神经元对维持觉醒至关重要。然而,很难想象演化会找不到一种重新设计大脑的方法,使神经递质补充无需关闭即可完成。大脑的某些区域在清醒和所有睡眠阶段都持续放电,所涉及的神经元能够无需离线即可补充资源。
生长激素和皮质醇(Growth hormone and cortisol)
除释放不同类型神经递质的神经元放电模式变化外,系统性激素的循环在睡眠期间也发生变化。其中,生长激素和皮质醇尤为重要,因为它们影响葡萄糖代谢。生长激素在入睡时增加,并在 NREM 的较深阶段达到峰值。另一方面,皮质醇水平在以 REM 睡眠为主的夜间后期阶段增加。与主要受昼夜节律控制的皮质醇释放不同,GH 的增加与睡眠启动相关(Van Cauter 等,1997117)。睡眠剥夺或在错误相位睡眠都因不同原因严重干扰葡萄糖代谢。这就是为什么健康睡眠对预防肥胖至关重要。
REM 稳态(REM Homeostasis)
与总体睡眠一样,REM 通过内稳态和昼夜节律过程控制。昼夜节律 REM 周期的峰值相位来得较晚,在主观夜晚后期。Benington 和 Heller 提出,是 NREM 睡眠的存在而非 REM 睡眠的缺失导致 REM 睡眠倾向的增加。慢波睡眠建立内稳态 REM 倾向,最佳的 REM 来自慢波”耗尽”与睡眠最后几小时出现的昼夜节律 REM 峰值的结合。学习与 REM 睡眠需求之间也存在强烈的内稳态联系。你学得越多,向 REM 的驱动越强。强化学习之后,REM 睡眠的分钟数和 REM 睡眠密度都会增加(De Koninck 等,1989118)。学习是直接影响 REM 需求还是经由 NREM 需求影响,目前尚不清楚。然而,有一点再清楚不过:重度学习者应该是重度睡眠者!
刺激基底前脑引起乙酰胆碱的释放,诱导清醒,同时也有利于 REM 睡眠。这意味着参与睡眠启动的基底前脑也参与 NREM/REM 转换。类似地,VLPO 细胞的一个子集有助于产生 REM 睡眠。
腺苷拮抗剂如咖啡因的影响也很重要。向基底前脑输注腺苷激动剂可增加 VLPO 中的 c-Fos,同时增加基底前脑释放的乙酰胆碱。已知乙酰胆碱诱导清醒和 REM 睡眠状态。由于激动剂输注,NREM 总量和 REM 睡眠总量都增加(Satoh 等,1999110;Scammell 等,200193)。
在其他内稳态激素影响中,睡眠中 VIP 和催乳素水平的升高促进 REM。物质滥用、延迟睡眠以及使用闹钟都可能导致 REM 睡眠亏欠(参见:REM 反弹假说)。
有一种睡眠理论认为 REM 睡眠帮助大脑从 NREM 睡眠中恢复,以加速清醒中的反应。这个理论通不过关闭检验,因为同样的恢复可能直接在清醒状态中进行,除非做梦中网络的辛勤工作由于某种未知原因是更快的恢复方法。然而,为什么大脑在完全”恢复”到清醒的条件下会经历 REM 反弹?关于组胺、5-羟色胺和去甲肾上腺素神经元需要恢复时间的说法听起来更合理,然而,它无法解释为什么我们需要具有不同神经递质、不同恢复策略的不同神经元群体。
向 REM 的过渡(Transition to REM)
在主观夜晚睡眠期间约一小时健康的 NREM 睡眠后,脑桥被盖中细胞的活动逐渐增加,该结构负责触发 REM 睡眠。负责触发 REM 睡眠的结构可能包括脚桥被盖核(PPN)和下外侧背侧被盖核(SLD)。GABA 能 SLD 和胆碱能 PPN 向多个方向发送信号。其中一个结果是肌肉张力丧失。另一个结果是丘脑、海马体和皮层的激活,并出现典型的 REM EEG。结果是大脑表现得仿佛内部醒了过来!在正在进行的 NREM 事件中向脑桥注射乙酰胆碱可触发 REM 睡眠,这说明了这种神经递质在睡眠周期调节中的重要性。在 REM 睡眠期间,皮层表现得如同清醒状态。在那个阶段体验到的梦境似乎是由脑干发送到皮层的随机脉冲产生的。皮层产生对该混沌输入尽可能最好、最连贯的意象。在梦境中我们体验到连贯的事件、真实的人、逼真的场景等。然而,所有这些都以最不可能的组合被拼凑在一起,仿佛大脑在测试各种”如果……会怎样”的场景。然而我们无法对梦境采取行动(患有暴力性睡眠的人除外)。负责 REM 控制的脑桥结构确保大脑输出被切断与移动肌肉的运动核团。我们常常在睡眠中想要行动(例如逃离一只凶猛的狗),却像陷在糖浆中一样纹丝不动。那时,只有眼睛快速移动,中耳的肌肉也抽动。
赋予 REM 其名称的眼球运动,由投射到上丘的脑桥核产生的脉冲控制。该脉冲与产生脑桥-膝状体-枕叶波(PGO)相关,PGO 波也用于检测 REM 睡眠。
在做梦期间,初级视觉皮层不活跃,而其次级区域活跃。这类似于人们被要求想象一个视觉场景而非实际看到该场景的情况。盲人的梦境更偏听觉和触觉。这似乎证实了 REM 睡眠在经验回放和记忆优化中的作用。他们在 REM 睡眠中也不表现出典型的眼动模式。这些观察导致了一种观点,即 REM 睡眠对创造力至关重要(比 NREM 睡眠更重要)。
REM 双稳态开关(REM flip-flop)
NREM 和 REM 睡眠之间的交互很可能由一个 REM 双稳态开关控制。一些科学家认为,在睡眠期间,中脑-脑桥交界处的一个超日节律振荡器控制着 NREM 和 REM 睡眠的规律交替。然而,考虑到交替的时机,双稳态开关模型是更好的类比。
与睡眠-清醒双稳态开关一样,REM 双稳态开关导致相对稳定的 NREM 和 REM 状态之间的持续切换,然而,该开关受各种内稳态输入的影响大得多,导致 NREM/REM 状态有些混乱的交替,在主观夜晚末期逐渐被 REM 昼夜节律峰值所主导。
多年来,胆碱能和单胺能状态之间的振荡似乎是对 REM 睡眠控制的最终答案。然而,一些不一致之处和新的研究导致了涉及 GABA 能结构的更新的类似模型(见下一节)。
REM 双稳态开关的模型(Models of REM flip-flop)
1962 年,Jouvet 表明刺激猫的尾侧中脑区或脑桥被盖产生类似 REM 睡眠的状态。这导致了一种假说:中脑-脑桥胆碱能结构负责 REM 睡眠中丘脑和皮层的激活。向脑桥网状结构注射胆碱能激动剂可增强 REM 睡眠这一事实也支持了该假说。
最初的相互作用模型——其中脑桥胺能和胆碱能神经元形成了一个经典的 REM-on/REM-off 双稳态开关——已被作为事实接受了四分之一世纪。新的研究已经鉴定出可能参与 REM 双稳态开关两侧的 GABA 能群体。
旧的 REM 双稳态开关包括在 REM(和清醒)中特别活跃的胆碱能 PPT 和 LDT,以及 BRF(脑干网状结构)。REM-off 组分由 DR(5-羟色胺)和 LC(NA)组成(Saper 等,2001119)。
较新的研究质疑了模型中的某些不一致之处。例如,对脑干胆碱能或单胺能核团的选择性损伤对 REM 睡眠的影响有限。REM-on 结构在 REM 期间低 c-Fos 表达也令人困扰。相反,有人提出关键的 REM-on 区域是 GABA 能 SLD(下外侧背侧被盖核)(Lu 等,2007120)。由于 SLD 不直接抑制 DR-LC,它们直接参与双稳态开关也受到质疑。SLD 的 GABA 能神经元投射到 vlPAG(腹外侧导水管周围灰质)和 LPT(外侧脑桥被盖),后者因此成为 REM-off 嫌疑。SLD 的损伤导致 REM 睡眠的丧失。
REM 双稳态开关的精确性质仍有待确定。新旧模型的组分也表现出一些相互作用。例如,PPT/LDT 确实兴奋 SLD 神经元,而 DR/LC 可能抑制 SLD 或激活 REM-off 组分。然而,这种相互作用并非直接相互的,因此旧组分被排除在新模型的核心之外。
睡眠的终止(Termination of sleep)
如果不是睡眠驱动的昼夜节律组分,睡眠控制系统将像一个无限的跷跷板。在睡眠接近尾声时,由视交叉上核(SCN)决定的昼夜节律困倦会产生睡眠倾向的下降,睡眠将在某次 REM 睡眠事件之后终止。正是 SCN 提供了最强授时因子——光——与昼夜节律周期之间的联系。SCN 内源性产生节律,但能够被光重置。来自视网膜的光脉冲传递到下丘脑和 SCN,产生停止与重置信号。睡眠结束将看到褪黑素释放的结束,取而代之的是另一种神经激素开始积累:5-羟色胺。一个假说说,正是高水平的 5-羟色胺让我们感受到了晨光带来的愉悦。高 5-羟色胺与警觉激素皮质醇结合,给我们一个良好的、警觉的新一天开端。除非你患有相位提前,否则一定要确保晨光能够照进你的卧室把你唤醒。
我们为什么需要睡眠?(Why do we need sleep?)
睡眠的生物学起源(Biological origins of sleep)
人类大脑是生物演化的最高成就,一切都始于简单的传导冲动能力,然后发展了神经网络这个惊人的概念。原始脊椎动物的大脑开始添加新结构以及优化神经连接丛林的新机制。睡眠是一个相对古老的发明,用于通过分子和神经机制重组记忆。昼夜节律在植物和不依赖于睡眠需求的动物中都已知。然而,演化 conveniently 把睡眠挂接到昼夜节律,以有效交替探索状态(即用大脑学习新事物)和巩固状态(即睡眠)。昼夜节律与约 100 种已知生理功能和参数相关,它们在一天中协同变化。与睡眠密切相关的是 5-羟色胺和褪黑素、ACTH和皮质醇、乙酰胆碱、腺苷和生长激素等激素水平的周期。
由于我们一生中三分之一的时间在睡眠,睡眠吸引了神经生理学家的大量关注,这毫不奇怪。鉴于大脑及其功能的巨大复杂性,有数百个理论试图解释睡眠的作用。只有最近,随着新研究技术的到来,我们才能在细节的海洋中看到大局。
睡眠理论(Sleep theories)
在过去两个世纪里,提出了几十个睡眠理论。一些科学家相信我们睡是为了记住,另一些相信我们睡是为了遗忘,还有一些相信睡眠与记忆无关。对生物学家来说,确定一个理论有效性的最佳基础是其演化视角。要问的主要问题是:睡眠服务于什么至关重要的功能,以至于需要关闭认知功能达一生三分之一的时间! 每个睡眠理论都必须通过这个主要检验,让我们称之为关闭检验(shutdown test)。Allan Rechtschaffen 说得最好:“如果睡眠不服务于绝对至关重要的功能,那么它是演化过程所犯的最大错误。” 关闭检验是消除不可信睡眠理论的最佳筛子。
通不过关闭检验的理论(Theories that fail the shutdown test)
以下是一些可能基于真实事实、真实模型或部分解释睡眠功能,但不解释每天”失去意识”数小时根本需要的理论:
- 节能(energy conservation):如果睡眠只是休息的一种形式,为什么我们不只是在黑暗的地方躺下,同时保持对捕食的警觉?大脑在夜间实际和清醒一样嗡嗡作响,所以关闭它不会提供任何额外好处,睡眠不是冬眠。
- 修复和恢复(repair and restoration):如果睡眠只是为了重建白天耗尽的身体,为什么不简单地用代谢逆转而不失去意识?许多组织可以在清醒时恢复和修复。
- 避免捕食(avoidance of predation):睡眠中的不活动本应最小化对捕食者的暴露,但为什么需要关闭所有感官并把反应时间延长到近乎无用的程度?而且肉食动物比杂食动物睡得更多,这暗示警觉可能比睡眠更能逃避捕食者。
- Siegel 理论:Jerome Siegel 博士提出自己的理论:“睡眠只是增加动物效率的好方法,例如只在狩猎条件好时狩猎,并最小化其风险。” 复杂睡眠控制机制的一小部分就足以满足 Siegel 提出的睡眠功能,例如,在适宜”睡眠”时间保持动物清醒和瘫痪(如 REM 睡眠)就足够了。
通过关闭检验的理论(Theories that pass the shutdown test)
- Krueger 和 Obal 理论:神经网络有进入睡眠状态的自然倾向。
- 梦境创造智慧:梦境是服务于记忆重组过程的表达。由于做梦,知识变得更连贯、更抽象、更适用。在睡眠中技能得到磨练和微调,这意味着我们早晨比睡前更聪明、更有技能!
- 垃圾回收(garbage collection):Evans(1965)的早期理论,源自计算机垃圾回收隐喻,说睡眠需要从无用记忆中清理大脑。确实,有大量关于睡眠中突触减弱的证据。
- 反向学习(reverse learning):Crick 和 Mitchison 1983 年在《Nature》上提出(Crick 和 Mitchison,1983110),人类记忆可能过载,REM 睡眠用于对记忆运行垃圾回收过程,称为反向学习,Crick 把那些累赘记忆称为”寄生记忆”,说*“REM 睡眠让我们能用更小的大脑服务于更大的目的”*。Crick 的开创性文章被许多研究者进一步发展,他们最终得出结论:REM 睡眠必须服务于由大脑神经网络突触权重表达的、记忆的优化!
- 记忆巩固(consolidation of memory):一些研究者假定记忆在 REM 睡眠中巩固。对于许多程序性任务,睡眠依赖的增强已被证明(Stickgold 和 Walker,2007111)。
- 激活-合成理论(activation-synthesis theory):Hobson 和 McCarley 1977 年提出(Hobson 和 McCarley,1977112),睡眠中存在感觉抑制,REM 期间存在运动抑制。脑桥 PGO 波上行到皮层,皮层试图解释那种刺激导致做梦。
- 大脑发育(brain development):在发育理论中,做梦本应为大脑回路提供内部刺激以确保生长和发育。REM 睡眠在发育视觉系统中的作用已被 Stryker 证明(Stryker 等,200110)。
睡眠作为神经优化器(Sleep as a neural optimizer)
上述通过关闭检验的所有理论,都与似乎是睡眠主要功能的内容兼容:
- 神经优化(neural optimization):一系列理论表明,睡眠是神经优化器。神经网络在一夜之间被重新布线,记忆从临时的、低容量、快速编码、快速关联、低干扰存储转移到可以安全地用于数月和数年而不受新编码记忆太多干扰的区域。抽象模式被提取,而遮蔽大局的细节被丢弃。更多见:Neural optimization in sleep。
睡眠与记忆(Sleep and memory)
NREM 与记忆(NREM and memory)
NREM 睡眠有魔力!如果你只睡几分钟并设法进入更深的睡眠阶段,你很可能醒来时感觉像全新的大脑。然而,这种短小睡只有在时机恰当且自然醒来时才可能。短时间 NREM 对学习的影响惊人:需要许多小时的繁重学习才能让大脑 homeostatically 困倦,而几分钟 NREM 睡眠就能带走那种困倦。
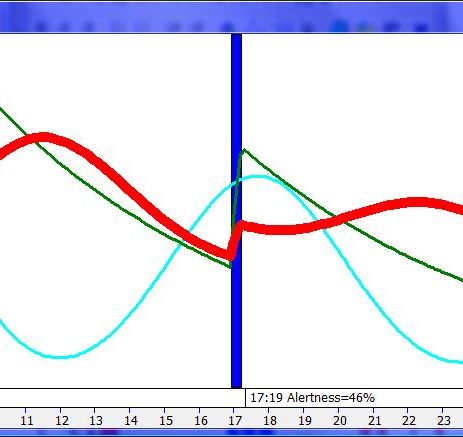
SuperMemo 中的双组分模型显示一次 19 分钟的小睡如何近乎翻倍内稳态警觉性组分(绿线)。(更多细节见:SleepChart 中的睡眠双组分模型)。这一理论模型有多年睡眠与学习数据支撑。
NREM 与海马体清理(NREM and hippocampal cleanup)
关于 NREM 睡眠功能最有说服力的假设,把它描绘成一个过程:存储在临时存储(主要是海马体、内嗅皮层及相邻结构)中的短期新记忆,被最优地写入皮层存储的庞大网络(Marshall 和 Born,2007113)。海马体连接大脑各区域以形成快速记忆关联,但容量有限,因此需要周期性清空,以便新的联结可以以更慢、更复杂的皮层存储方式记录。假设这发生在 NREM 睡眠中,且相当快。科学家早就注意到,在慢波睡眠中,海马体和新皮层网络倾向于重放与新经验相关的放电模式(Wilson 和 McNaughton,1994114)。
NREM 与陈述性记忆(NREM and declarative memory)
慢波睡眠剥夺对陈述性记忆的影响大于程序性记忆(Plihal 和 Born,1997108)。换言之,考前削减睡眠有效地使保留为考试所学的知识变得更难。这种效应在长期尤其明显,而在考试当天不那么明显,这就是为什么这么多学生一遍又一遍犯同样的错误——他们在最后一夜换取更多学习时间,代价是所学知识的长期保持。
尖波爆发与长期记忆(Sharp wave bursts and long-term memory)
在深睡中,可以在海马体记录到 SPW 爆发(尖波爆发)。一些研究者认为,这可能是记忆巩固的关键时刻,海马体充当新皮层的神经训练器,长期记忆存储在跨皮层联结中。在 SPW 爆发期间,一天的经验将被最优地通过神经训练转移到新皮层网络。随后将启动基因表达和蛋白质合成,这两个过程都是修改长期突触权重所必需的。蛋白质合成构成了将持续数月乃至数年的记忆的开端(如果通过重复/复习来维持,例如用 SuperMemo)。更多细节参见 Molecular correlates of the two-component model of long-term memory(Wozniak 等,1998121)。这些长期记忆不进入睡眠周期的适当阶段就无法形成!没有睡眠就无法建立长期记忆。此外,如果早晨被闹钟截断,学习的效率也会降低。
用干扰测试检测记忆优化(Detecting memory optimization with interference tests)
所有关于陈述性记忆的研究都可能受到睡眠中发生的神经优化的混淆。睡眠通常具有重构的形式,其中相同的记忆以不同方式存储。这样,记忆改变的效果可能无法看到,但其内部表征会变化。这种重构可能无法用行为测试检测,或非常难以测试。同一个法语单词存储在工作记忆中与存储在长期存储中感觉是一样的。它似乎不会在一夜之间变异,即使会,变化也很难察觉。然而,睡眠不足会影响存储不完美的记忆,多于那些存储已优化的记忆。研究者们在采用干扰技术展示睡眠如何防止记忆被新信息覆盖之前,一直难以确认睡眠对陈述性记忆的重要性(Ellenbogen 等,2006122)。使用 SuperMemo,有可能收集使得类似的长期测定成为可能的睡眠数据。由于缺乏明确的睡眠限制数据,目前我们只知道记忆巩固随清醒日的推进而逐渐变差(参见:记忆巩固)。
REM 与记忆(REM and memory)
REM 睡眠的功能有许多理论。REM 睡眠与学习相关已被多个实验发现,然而一些科学家质疑这些发现,指出实验错误或抗抑郁药是强 REM 抑制剂却不损害记忆的事实。今天我们知道,不间断 NREM 和 REM 睡眠自然、和谐的交互对记忆、学习和创造力至关重要(Salzarulo 等,2000115)。
REM 与学习(REM and learning)
Carlyle Smith 1991 年表明,在大鼠的 REM 睡眠窗口期间给予蛋白质合成抑制剂,会阻止睡眠后通常出现的行为改善。这是 REM 睡眠对记忆重要的有力指示。此外,程序性学习的增加伴随 REM 密度的增加,且增加程度与个体的学习能力成正比(Smith 等,200487)。
REM 作为一种神经训练(REM as a form of neural training)
一个假设说,REM 睡眠是大脑的一种训练形式。正常清醒活动用新的活动模式训练海马体,REM 睡眠做同样的事,只是使用想象的、随机化的假设模式。仿佛大脑在清醒中得不到足够,它需要在睡眠中更多特殊训练。这种额外训练会很有益,因为它成本很低(不需要消耗行为能量),并利用已存储在大脑中的信息。
REM 与创造力(REM and creativity)
随着夜间进展,REM 增多,NREM 减少。如果”REM 训练”假设正确,最后以 REM 睡眠结束的睡眠段可能为早晨大脑提供下一个大想法,这可能是科学史上丰富的创造性早晨洞见的构建块。Walker 在不同睡眠阶段醒来的受试者中研究字谜表现时收集了一些支持这一观念的证据(Walker 等,2002116)。如果关于最后一个 REM 片段创造性贡献的推理正确,我们可以得出一个戏剧性结论:**闹钟可能是现代世界大想法的主要杀手!**压力或激烈竞争也 guilty 于破坏人类创造力,但在一个没有足够睡眠的无压力世界里控制压力以获得好睡眠,比产生伟大想法要容易。研究者和教育者在淡化闹钟造成的损害时应非常谨慎(参见:Jim Horne 和 Daniel Kripke)。父母也应对那些无法按时醒来上学的孩子表现出更多宽容(参见:睡眠与学校)。
REM 作为神经优化器(REM as a neural optimizer)
如果 REM 睡眠只是睡眠中的一种训练或创造性选项,为什么我们会产生 REM 亏欠?昼夜节律 REM 倾向可能提供了 NREM 与 REM 之间的平衡。内稳态 NREM 压力可能源于学习(在清醒或 REM 中)。然而,内稳态 REM 睡眠倾向以及 REM 剥夺后的 REM 睡眠反弹都似乎表明,REM 远不止是一个选项。REM 剥夺对复杂任务和程序性记忆的复杂影响,可能是 REM 睡眠更重要功能的表达,其中神经优化基于与 NREM 睡眠相比大脑中信息流方向的反转。NREM 可能仅充当长期记忆存储工具,而没有太多优化网络布局的能力。NREM 可能只是一种抗干扰工具。然而,睡眠帮助组织记忆、增加其抽象性并降低存储成本。也许这种优化作用完全由 REM 睡眠承担。也许伪模式训练使得将广泛的、昂贵的网络记忆迁移到更小、更通化的回路成为可能(参见:睡眠中的神经优化)。一个重要线索来自婴儿睡眠。由于新生儿探索其世界的能力有限,REM 睡眠的探索功能可能在发育中扮演重要角色。然而,Siegel 博士注意到,人类在 REM 中花费的时间与其学习能力并无良好相关。我相信,这种不一致可以用个体差异来很好地解释,这些差异不仅表现在学习能力或每晚 REM 的平均量上,还表现在 REM 的效率(即学习-REM 比)上。这类似于,例如,我们的消化能力。有些人可以大吃大喝然后全部排出去。另一些人吃得很少,却仍能将所有营养提取到最后一毫克(并胀气)。如果我们进一步观察各个物种,我们会发现 REM 量与”动物智商”之间的联系更弱。这可以解释为:聪明的动物会从 REM 睡眠中提取多得多的价值。换言之,无论多少优化都无法在容量小的网络中创造奇迹。更令人困扰的可能是这一主张:在鲸类(尤其是幼崽)中很难检测到任何 REM(Castellini,2002123)。如果 REM 真如本章所描绘的那样对程序性学习不可或缺,那么它在年轻聪明的掠食性游泳者身上似乎也是不可或缺的。显然,作为一个相对较新的演化实体,基于 REM 的神经优化可能有其变体,具有不同的现象学,可能不会立即被研究者发现。此外,以单半球慢波睡眠奇迹为特征的鲸类睡眠,有超过 5000 万年的时间来发展出使其区别于人类 REM 睡眠的特性——这占了 REM 睡眠存在的很大比例。
突触在睡眠中的变化(Synaptic changes in sleep)
一些研究表明突触在睡眠中得到强化,而另一些研究发现了相反的效果。清醒期间整体突触强度倾向于增加124,而学习能力持续下降。清醒增加所有行为状态下的皮层放电频率(Tononi 等,2009125)。睡眠中选定突触同时发生的减弱和强化最好用某种一夜之间发生的记忆重组来解释。这并不与睡眠的记忆巩固功能矛盾,也与清醒期间学习能力倾向于下降这一事实不矛盾。
在研究睡眠中突触强度的变化时,我们始终需要区分:
- 短期记忆:一天学习导致的突触导电性的短期增加
- 长期记忆:回忆旧记忆的能力(例如用 SuperMemo 测量)
打个比方,大脑就像一台白天不断向其内存加载数据块的计算机(短期记忆)。随着内存填满,计算机变慢,所有应用程序都陷入停滞。然而,如果你测试单个内存单元,你会发现它们紧紧抓住新数据。在夜间,计算机将逐渐组织这些数据块,消除差异和重复,将记忆写入硬盘(长期记忆),并运行碎片整理以便于快速访问。我们需要审视这个比喻的神经生理学关联,以及清醒期间回忆减弱的最可能解释,因为清醒期间突触导电性的增加和白天学习能力的下降都有充分记录。最连贯、最有吸引力且最有支持的假说说,短期低干扰网络的过载是负责清醒日期间记忆能力下降的原因(参见:NREM 与记忆)。这种下降使工作记忆瘫痪,进而影响人类认知能力的整个谱系。睡眠的主要功能就是重新分配、重新巩固并优化那些拖慢进一步学习的短期记忆。
至于睡眠中突触强度的下降,它也很符合目前的睡眠和学习模型。睡眠的主要功能之一应该是优化记忆存储。这需要以最有效的方式表征记忆,使它们最抽象、占用最少空间、产生最小干扰等。该过程确实应导致降低记忆的整体成本,并导致冗余突触连接的减弱。
Tononi 博士认为,清醒活动产生整体突触权重的增加,睡眠可能需要平衡这种增加。假设的下调将发生在慢波睡眠中(Tononi 和 Cirelli,2006126)。Tononi 博士将记忆层级中从短期(磷酸化)到长期(AMPA 运输)到重塑(出芽)的不同组分聚集在一起,而我宁愿把赌注押在日常学习和短期记忆上。整体皮层下调可以是学习后清理的优美表达,与 Crick 和 Mitchison 的观点一致。这种清理可以与由短期记忆结构(如海马体、杏仁核等)主导的选择性突触强化相结合。
睡眠中的神经优化(Neural optimization in sleep)
海马体损伤(Hippocampal lesions)
著名的 H.M. 病例(双侧海马体切除)表明,海马体对形成新的陈述性记忆至关重要,但术前记忆完好。这符合海马体作为临时存储、新皮层作为长期存储的模型(Squire 等,1984127)。
时间等级性逆行性遗忘(Temporally graded retrograde amnesia)
海马体损伤的患者,近期记忆受影响最大,而遥远的旧记忆完好。这表明记忆随时间从海马体转移到新皮层,这个过程发生在睡眠中(McClelland 等,1995128)。
睡眠中的记忆处理(Memory processing in sleep)
睡眠研究的两大主线是记忆巩固和神经优化。记忆巩固是指新获得的记忆在睡眠中被稳定下来,使其更不容易被遗忘。神经优化则更进一步,指的是大脑在睡眠中重新组织记忆,使其更抽象、更高效、更不容易与新信息产生干扰。这两条研究主线正在汇合,共同指向睡眠作为大脑”夜间优化器”的核心角色。
灾难性遗忘(Catastrophic forgetting)
人工神经网络在用新数据连续训练时会灾难性地遗忘旧数据,这是人工神经网络的主要局限之一。人脑如何避免灾难性遗忘?答案在于互补学习系统:海马体作为快速、低干扰的临时存储,新皮层作为慢速、高干扰的长期存储(McClelland 等,1995128)。
睡眠中的两阶段记忆处理(Two-stage memory processing in sleep)
Buzsáki 的两阶段模型是理解睡眠中记忆处理的核心(Buzsáki,19894;Buzsáki,1996129):
- 第一阶段(清醒/探索,theta 节律):海马体快速编码新经验。
- 第二阶段(睡眠,尖波涟波):海马体重放清醒期间的模式,训练新皮层形成长期记忆。
这个过程在睡眠中反复进行,海马体的尖波涟波(sharp wave ripples)是新记忆从海马体转移到新皮层的关键机制(Wilson 和 McNaughton,1994114;Lee 和 Wilson,2002130)。互补学习系统理论为这个两阶段处理提供了理论基础:如果海马体和新皮层使用相同的学习速率,系统要么无法快速学习新事物,要么会因为新学习而灾难性地遗忘旧知识。海马体的高速学习与新皮层的慢速优化之间的分工,正是避免这一困境的关键。
优化记忆(Optimizing memories)
神经优化理论的核心假设是:睡眠不只是巩固记忆,而是优化记忆。这意味着记忆在睡眠中会变得:(1)更抽象——提取共性、丢弃遮蔽大局的细节;(2)更高效——以更少的突触资源存储;(3)更少干扰——不同记忆之间的相互干扰降低。这一过程类似于程序员对代码进行的”重构”(refactoring):代码的功能不变,但其内部组织变得更优雅、更高效。同样,一个法语单词在你睡前和睡后感觉一样,但其在大脑中的存储方式已经发生了深刻变化。
垃圾回收(Garbage collection)
受计算机垃圾回收隐喻的启发,Evans(1965)和后来的 Crick 与 Mitchison(1983)提出,睡眠的一个功能是清除大脑中无用的、矛盾的或”寄生性”的记忆。在计算机科学中,垃圾回收是指自动释放不再需要的内存以供重用的过程。在大脑中,这一过程表现为睡眠中冗余突触连接的减弱和无用记忆痕迹的清除。这与 Tononi 和 Cirelli 的突触稳态假说一致:睡眠中的整体突触下调可被视为一种全局的”垃圾回收”,为第二天的新学习腾出空间。
单半球睡眠(Unihemispheric sleep)
一些海洋哺乳动物(如海豚和鲸类)和鸟类表现出单半球慢波睡眠——一个半球在睡眠时,另一个半球保持清醒。这种非凡的适应使它们能够在睡眠时保持游泳或警惕捕食者。单半球睡眠的发现对睡眠理论提出了有趣的挑战:如果睡眠的主要功能是整体性的神经优化,那么单半球睡眠如何实现这一功能?一个可能的解释是,单半球睡眠动物可能在两个半球之间交替进行优化,尽管这种优化的效率可能不如双半球同时睡眠。有趣的是,在鲸类(尤其是幼崽)中很难检测到任何 REM(Castellini,2002123),这进一步说明睡眠的形式和功能在演化中可能有显著的变异。
睡眠中的问题解决(Problem solving in sleep)
许多创造性的”灵光一闪”发生在睡眠之后。Wagner 等(2004)研究表明,睡眠激发洞见(Wagner 等,2004131)。大脑在睡眠中重组网络,能在相距甚远的神经元节点间建立创意联结,这就是为什么许多棘手的科学或编程问题往往在睡了一觉后迎刃而解。
结论(Conclusions)
睡眠的神经优化理论综合了大量看似矛盾的研究发现。睡眠不只是休息,不只是巩固,而是对神经网络进行深度优化的过程。在这个框架下,NREM 主要负责将记忆从海马体的临时存储转移到新皮层的长期存储,而 REM 则负责对这些记忆进行创造性重组和优化。两种睡眠阶段的和谐交替对记忆、学习和创造力至关重要。这一理论也为睡眠卫生提供了坚实的理论基础:尊重睡眠就是尊重大脑自我优化的过程,而任何干扰这一过程的因素(如闹钟、睡眠剥夺、物质滥用)都可能损害我们的智力潜能。
并非所有科学家都同意(Not all scientists agree)
2001 年 11 月 2 日的《Science》包含了关于睡眠生理学的对立文章,为关于睡眠功能的学术争端提供了更多燃料(Siegel,2001132;Stickgold 等,2001133;Maquet,2001134)。当解开被称为睡眠的复杂神经优化过程细节的努力接近带来重大突破时,有声誉的研究者却从中作梗。著名的发作性睡病专家 Jerome M. Siegel 博士分析了睡眠作用的证据,发现证据 scant!
Robert Vertes 和 Jerome Siegel
Jerome Siegel 不是唯一质疑睡眠在记忆中作用的异见者,Robert Vertes 也是高度有声望的睡眠研究者。Siegel 和 Vertes 来自美国东西两岸的不同领域,却联合起来对抗关于睡眠参与处理记忆的高度既定观点。2005 年,他们在《Sleep》上发出呼吁:Time for the Sleep Community to Take a Critical Look at the Purported Role of Sleep in Memory Processing(Vertes 和 Siegel,2005135)。
1. 睡眠不服务于陈述性记忆?
研究在这方面似乎相当矛盾。一天所学词汇的回忆,似乎不受次夜缺乏睡眠的太大影响(除了睡眠剥夺后的日子)。然而,用 SuperMemo 获得的数据明确显示,睡眠是为人脑记忆新输入”拔插头”的显著工具!
2. REM 睡眠剥夺不导致认知损伤?
Vertes 和 Siegel 写道:“几条证据表明 REM 睡眠不参与记忆处理/巩固。最强有力的论据或许是,脑干损伤个体或服用抗抑郁药个体中 REM 睡眠的显著抑制或消除,对认知没有不利影响。一个经典案例是一位以色列男子,19 岁时遭受脑干损伤,33 岁检查时显示无 REM 睡眠。该男子现年 55 岁,是律师、画家,有趣的是还是一家以色列杂志的谜题专栏编辑。最近评论他那位’著名’病人时,Peretz Lavie 说’他可能是我认识的最正常的人之一,也是最成功的人之一’。”
然而,这不能用于贬低 REM 的重要性。首先,REM 的表达是复杂和多线程的,我们不知道损伤的是 REM 控制系统的哪部分。其次,过”正常生活”是认知评估的非常弱的测试。再次,大脑的代偿能力巨大。根据 Buzsáki,REM 睡眠可能是帮助从新皮层向海马体回路加载模式(用于后续 NREM 睡眠中记忆重写)的主要优化器,如果考虑这种解读,REM 剥夺允许”正常”生活,但一个人的创造性洞见会显著减弱。
3. 睡眠依赖的程序性学习增强未被证明?
理论上,由于其随机性质,程序性学习应比陈述性学习更受益于睡眠。Matthew Walker 博士的研究相当有说服力。有趣的是,Siegel 自己 2001 年曾告诉 WebMD:“程序性学习的研究清楚表明,在你睡眠之前不会得到改善。“
4. 清醒中的学习远比夜间增强重要?
overnight 对学习效果的改善,不比繁重锻炼后慵懒睡眠中的肌肉增长更违反直觉。比较清醒中学习过程的价值与夜间优化的价值,与量化大脑和心脏对我们生存的贡献没什么不同。两者都至关重要且不可或缺。
5. 睡眠模型应该简单
如 always,lex parsimoniae(奥卡姆剃刀)是科学的有益指导。睡眠作为神经优化器的概念似乎符合令人信服的简单性标准。
REM 中的随机脉冲如何使梦境有意义?(How can random impulsations in REM make a sense in dreams?)
Vertes、Siegel 和 Hobson 等怀疑者指出,如果在 REM 中,脑桥向皮层发送的脉冲是随机的,那么从中产生的梦境如何能服务于记忆的优化?答案来自神经网络理论。即使是随机的输入,当作用于一个已经存储了有意义模式的神经网络时,也会产生有意义的输出。梦境中看似荒谬的场景,实际上是大脑利用已有知识对随机刺激进行解释的结果。这个过程本身就是一种优化:它测试网络的稳健性,发现并修复薄弱环节,并在看似无关的记忆之间建立新的联系。这就是为什么梦境既荒谬又深刻——它是大脑在做”如果……会怎样”的思想实验。
Siegel 博士的睡眠理论
以下是 Siegel 博士几个与我关于睡眠生理学和演化的知识不太吻合的陈述:
- “睡眠只是增加动物效率的好方法,例如只在狩猎条件好时狩猎,并最小化其风险。” 对于那个特定表述,复杂睡眠控制机制的一小部分就足以满足 Siegel 提出的睡眠功能。
- “我的理论并不意味着睡眠中不发生重要的事——它只意味着睡眠没有至关重要的通用功能。” 演化告诉我们,如果一个功能在数百万年间保守,那几乎保证存在潜在的至关重要的组件。正是神经优化在数百万年间保留睡眠作为动物生活不可避免的一部分。
我个人的偏见(My personal bias)
我提到过我个人的偏见。我生命中有一件事让我困惑并留下持久印记。那大约是 1980 年,我的共产主义制造的收录机坏了,我离不开重口味黑人放克,所以拼命想修收录机。然而尝试半天后我束手无策,极度失望地去睡觉。当我早晨醒来,我意识到我可能梦到了修理,甚至模糊记得一个让收录机复活的修理技巧。当我头脑清醒后,我确信我在睡眠中发现了修理方法,起床第一件事就是试那个方法——收录机复活了!那天,我变得认知偏向于相信睡眠在我们大脑中产生新质量!多年来,我不断注意到类似的 overnight 突破(在一天失败的问题解决之后)。我不断确认,早晨的大脑与傍晚的大脑处于完全不同的状态。是的,我有偏见,凭借所有那些轶事证据,我毫不怀疑大脑在睡眠中进行繁重的记忆处理(另见:Sleep inspires insight(Wagner 等,2004131))。
橄榄枝(Olive branch)
Matthew Walker 2009 年做出如下预测:“我们很快将有一种新的、睡眠依赖记忆处理的分类法,它将超越过去两极化的全有或全无观点。有了这些发现,我们可以重新欣赏清醒和睡眠如何以共生联盟结合,协调我们记忆的编码、巩固和整合,其最终目标可能是创建一个不依赖于冗长保留所有先前所学事实的、存储知识的通用目录。“(Walker,2009136)
给怀疑者的更多阅读(More reading for skeptics)
如果你对 Vertes 和 Siegel 博士的观点感兴趣,见:
- Siegel 博士的大量有趣出版物:http://www.semel.ucla.edu/sleepresearch
- Robert Vertes 博士 2000 年:“我们相信累积证据表明REM 睡眠在记忆处理或巩固中不起作用](http://web.archive.org/web/20100113034337/http://bbsonline.org/Preprints/OldArchive/bbs.vertes.html)“
生物钟基因(Clock genes)
生物钟的分子机制基于一组生物钟基因的转录-翻译反馈环路。在哺乳动物中,核心生物钟基因包括 CLOCK、BMAL1、PER(PER1、PER2、PER3)和 CRY(CRY1、CRY2)。这些基因的蛋白质产物形成反馈环路,产生约 24 小时的分子振荡。这些基因的突变可以改变昼夜节律周期的长度,导致睡眠相位障碍(如家族性 ASPS)。有趣的是,在一些蓝藻中,生物钟可以在没有转录-翻译反馈的情况下运行(Tomita 等,2005137;Qin 等,2010138),这表明存在转录后水平的生物钟机制。这些发现对我们理解生物钟的基本原理有重要意义。
迷思与事实(Myths and facts)
睡眠长度(Length of sleep)
- 迷思:我们每晚应该睡 8 小时。事实: 许多人可以睡得更少,他们不需要追求 8 小时;另一些人可能需要超过 8 小时。我们都应恰好睡我们睡眠系统所要求的那么多小时。试图获得规定的 8 小时睡眠会驱使一些人走向失眠,而另一些人则仍会睡眠剥夺。
- 迷思:睡得少的人活得更长。 2002 年,Kripke 博士比较了睡眠长度与长寿,发现睡 6-7 小时的人比睡 8 小时及以上的人活得更长。事实: 最好的长寿前景是通过符合自己自然身体节律的睡眠来确保。坚持自己良好节律的人常常睡得更少,因为他们的睡眠结构更好(因此更令人神清气爽)。“自然睡眠”的人活得更长,那些违背身体呼唤睡觉的人常常需要更多小时仍感觉不神清气爽。此外,疾病常与增加的睡眠需求相关,相关不等于因果。
- 迷思:如果你困倦,意味着你没睡够。事实: 对神清气爽的头脑来说,有比睡眠长度更重要的因素。同一个人在不同情况下,执行良好的 3 小时睡眠后可能比一长夜低质量睡眠后更不困倦。导致白天困倦的主要因素是:使用闹钟、在错误相位睡眠(过早或过晚)、睡眠呼吸暂停、用安眠药或酒精诱导睡眠以及物质滥用。
- 迷思:身体总会像渴望过量食物一样渴望过量睡眠。事实: 与脂肪储存不同,额外睡眠似乎几乎没有演化益处。一旦他们补上并进入节律,睡眠长度实际上很可能下降。
- 迷思:少睡让你更有竞争力。事实: 长期最大竞争力的唯一好公式是通过最大健康和最大创造力。你只有在完美命中昼夜节律低谷时才能获得最短、最高质量的睡眠。错误时段的睡眠或被闹钟打断的睡眠注定削弱你的智力表现和创造力。
- 迷思:我们可以每天睡 3 小时。事实: 如果你的健康不是考虑因素、智力能力不是关键,这或许可行。如果你的抱负超越于此,你应该恰好睡身体想要的那么多,这才是聪明人的最佳选择。
- 迷思:你可以提前积累睡眠好处。事实: 如果你预期一个不眠之夜,自然最好是状态良好,前一晚的好睡眠会有帮助。然而,你不会从前一周或一个月的良好睡眠中获得太多好处。睡眠不是食物,你不能为未来使用提前积累。睡眠信用不存在。
打盹(Napping)
- 迷思:避免小睡。事实: 小睡确实可能加重 DSPS 患者的失眠,尤其是当天晚些时候进行时。否则,小睡对智力表现高度有益。
- 迷思:小睡是软弱的标志。事实: 小睡不是软弱、不健康、懒惰或缺乏活力的标志,它是双相睡眠节律的系统发生残留。
- 迷思:小睡降低预期寿命。事实: 习惯性小睡延长寿命。
- 迷思:小睡可能导致糖尿病。事实: 中国一项研究发现小睡与糖尿病相关(Chen 等,2010121),但它没有区分小睡类型或暗示因果。时机恰当的小睡应降低糖尿病、心血管疾病等风险。
- 迷思:小睡前的咖啡有帮助。事实: 咖啡可能帮助你从小睡中快速醒来并感觉相当神清气爽,然而如果咖啡因阻止小睡走完自然进程,那种清新可能更快消散。只在赶时间时(如长途驾驶中途停车)在小睡前喝咖啡。
- 迷思:小睡总是好的。事实: 时机不当的小睡可能加重失眠、移动睡眠相位或导致睡眠惰性。
- 迷思:长小睡让你进入深睡并导致睡眠惰性。事实: 时机恰当的小睡(通常在自然醒来后第 7 小时左右)越长,警觉性和学习能力的提升越大。
- 迷思:百灵鸟应在下午 1 点小睡,猫头鹰应在下午 2:30 小睡。事实: 百灵鸟和猫头鹰在偏好小睡时机上差异很小,都大约在自然醒来后 7 小时。
- 迷思:每个人都有自己最佳的小睡长度。事实: 最佳小睡长度不存在!小睡受许多内稳态调节输入影响,长度会因情况而日复一日变化。
- 迷思:20 分钟的能量小睡最有效。事实: 时机恰当的小睡自然终止时最有效,可能持续 3 分钟或 3 小时。
- 迷思:不要小睡,你只会更累。事实: 时机恰当的小睡可以使你的生产力翻倍。
- 迷思:用闹钟把小睡保持在 15 分钟,从第 2 阶段 NREM 醒来以避免昏沉。事实: 时机恰当的小睡在健康昼夜节律周期中永远不应让你昏沉。
- 迷思:失眠者应避免小睡。事实: 早小睡和习惯性小睡者的小睡对夜间入睡影响可忽略。
睡眠惰性(Sleep inertia)
- 迷思:大多数人早晨经历睡眠惰性。事实: 早晨的睡眠惰性完全可以避免,扔掉闹钟后你不太可能经历睡眠惰性。
- 迷思:醒来是一个需要自行进展的缓慢过程。事实: 在正确时间没有闹钟帮助醒来应该相当快,好日子 3-5 分钟。
- 迷思:过量睡眠导致头痛和昏沉,较短的睡眠更健康。事实: 这是又一个因果混淆的例子。睡眠剥夺的人常常睡得很长以偿还睡眠债,长恢复睡眠确实常导致头痛,但问题首先是短睡眠造成的。
- 迷思:睡眠惰性不显示昼夜节律周期。事实: 取决于睡眠惰性类型,纯昼夜节律的错误相位惰性会随昼夜节律低谷过去自行消失。
- 迷思:小睡导致睡眠惰性。事实: 只有太晚的小睡导致睡眠惰性,时机恰当的小睡应相对短(30-90 分钟)且非常令人神清气爽。
- 迷思:睡眠惰性与先前睡眠持续时间相关。事实: 睡眠惰性由破坏自然睡眠过程造成,与先前睡眠长度关系不大。
- 迷思:增加大脑血流是睡眠惰性的好疗法。事实: 只有健康的睡眠才是睡眠惰性的真正疗法,咖啡因、运动、噪音、强光、压力等只能掩盖它。
失眠(Insomnia)
- 迷思:安静和黑暗对睡眠至关重要。事实: 安静和黑暗确实使入睡更容易,但它们并非至关重要。对数百万失眠者来说,专注于宁静的睡眠场所遮蔽了大局:让我们睡好的最重要因素(假设健康良好)是遵守自己的自然昼夜节律!按自然节律入睡的人常能在明亮的阳光下睡好。
- 迷思:咖啡因可导致失眠。事实: 咖啡因可使失眠恶化,但从来不是主要原因。
- 迷思:失眠由缺镁引起。事实: 失眠有多种原因,矿物质缺乏甚至排不到原因清单的前列。失眠最常见的原因是压力、健康不佳和在错误相位(过早)睡眠。
- 迷思:分段睡眠是自然的。事实: 分段睡眠可能是”过量夜间”的表达,可见于人们过早入睡的现代条件。
- 迷思:镁、叶酸和其他补充剂可以帮助你睡得更好。事实: 健康所需的营养也对睡眠有益,然而补充不太可能在解决睡眠问题上起显著作用。
就寝时间(Bedtime)
- 迷思:每天同一时间上床对你有好处。事实: 严重 DSPS 患者可能根本无法每天同一时间入睡,强制尝试只会导致压力和失眠的自我喂养循环。
- 迷思:午夜前的睡眠更有价值。事实: 睡眠在你自己生物钟机制计划的时间到来时最有价值。
- 迷思:最好与太阳一同醒来。事实: 你应在身体决定睡够时醒来。
- 迷思:早睡早起使人富有、健康和明智(本杰明·富兰克林)。事实: 许多百岁老人是早起者,但早起者只有在早起习惯自然时才长寿。如果你的主要工作工具是大脑,不要遵循这个建议!只在自然时醒来,即便你的自然醒来时间晚。
- 迷思:早睡让我们更有精力。事实: 最大早晨清新和能量来自在正确相位睡眠。
生物钟(Body clock)
- 迷思:人体生物钟运行 24 小时周期。事实: 周期实际上长于 24 小时,大脑使用重置按钮把周期与活动模式同步。
- 迷思:人是早晨型或夜晚型。事实: 这更多是误称而非迷思。人们真正不同的是生物钟周期以及对重置该节律的刺激的敏感性和可得性。
- 迷思:你无法改变生物钟固有的周期长度。事实: 通过各种时间疗法技巧,可以略微改变时钟周期。
- 迷思:人体生物钟可以轻松适应各种作息。事实: 时钟周期长度只能略微改变(也许最多一小时),每天也只能略微推动相位(也许最多 2 小时)。
- 迷思:夜班不健康。事实: 有害的不是夜班,而是睡眠节律从白天到夜间及反向的不断切换。
儿童睡眠(Sleep in children)
- 迷思:新生儿几乎整天都在睡。事实: 新生儿更可能花掉他们一半的日子睡眠,其中一半是 REM(相对于通常成人的 20%)。
- 迷思:婴儿睡前不应玩耍。事实: 除非玩耍极其令人兴奋或以任何方式有压力,否则玩耍增加睡眠的内稳态压力,应使婴儿更容易入睡。
- 迷思:上学迟到是坏事。事实: 持续无法醒来上学的孩子应被放任,他们神清气爽的头脑和健康远更重要。
- 迷思:上学迟到是懒惰的标志。事实: 患有 DSPS 的年轻人可能持续有问题按时起床上学,这些孩子常常实际上比平均更聪明,绝不是懒惰。
学习与创造力(Learning and creativity)
- 迷思:天才睡得少!事实: 看爱迪生和特斯拉,很容易相信减少睡眠对创造性成就似乎不构成问题。然而在睡眠剥夺条件下尝试创造性工作的人会很快发现,神清气爽的头脑远比少睡省下的 2-3 小时重要。天才睡得少的说法不真实,例如爱因斯坦承认他如果获得扎实的 9 小时睡眠区块工作得最好。
- 迷思:学习前的睡眠会削弱学习效果。事实: 时机不当的睡眠可能导致睡眠惰性,在睡眠惰性条件下学习特别低效。然而睡眠实际上是学习的最佳盟友,只要睡眠在正确相位、不被打断、不被物质或睡眠药物削减,最佳学习发生在早晨。
- 迷思:既然我们睡后感觉休息好了,睡眠一定是为了休息。事实: 睡眠是为了优化记忆。尽管看似不活动,睡眠中的代谢率只下降 15%,平均一夜的节省相当于一个苹果所含能量。为有效编码记忆,哺乳动物、鸟类甚至爬行动物需要关闭思维并在大脑中做些家务,这对生存至关重要。
警觉性(Alertness)
- 迷思:我们早晨 8 点最不警觉,警觉性在白天增加。事实: 警觉性在一天中有两个峰值。对大多数健康睡眠作息的人来说,警觉性在醒来后 30-80 分钟最高。
- 迷思:青少年在傍晚显示更强警觉。事实: 青少年有协调晚间活动与早晨学校作息的自然困难,他们的傍晚警觉来自主要由生活方式诱发的延迟相位移问题。
闹钟(Alarm clock)
- 迷思:闹钟可以帮助调节睡眠节律。事实: 闹钟可以帮你把睡眠节律推入期望框架,但很少帮你实现健康睡眠节律。实现健康睡眠和健康睡眠节律的唯一可靠方法是:只有真正困倦时才入睡,自然醒来没有外部干预。
安眠药(Sleeping pills)
- 迷思:安眠药可以帮助你睡得更好。事实: 安眠药可以帮助你入睡,但这种睡眠质量远低于自然诱导的睡眠。在睡眠医学上至关重要且无法用其他方式实现时,安眠药可能有用,否则只要可能就避免。
- 迷思:褪黑素可用于提升小睡。事实: 恰恰相反,褪黑素可能让小睡感觉非常不令人神清气爽。
多相睡眠(Polyphasic sleep)
- 迷思:我们可以适应多相睡眠。事实: 我们基本上是双相的,所有改变内置节律的尝试都会导致健康、时间和脑力的损失。简单规则是:困了就睡;睡着了就不间断继续。参见:多相睡眠的迷思
增量写作(Incremental writing)
本文采用一种名为**增量写作(incremental writing)**的创作写作技巧写成。
SuperMemo 可用于创作写作过程,它将文本写作与增量阅读(incremental reading)过程的学习和复习结合起来。类比增量阅读,这一过程被称为增量写作。增量阅读与增量写作的主要区别在于,文章的”全局图景”是在集合(collection)中构建的,而非在作者脑中。这适合难以按顺序组织的大型、事实密集的材料。此外,作者自己的写作可能是大部分输入的来源,而非外部电子来源。增量写作也适合对大量先前写作的汇编,尤其是重复性、事实丰富且常常松散关联的材料。对于线性思路的文本,增量写作用处较小。本文是 supermemo.com 上与睡眠和学习相关的多份文本的汇编。所有相关材料都被导入 SuperMemo 15 并以增量方式处理。文章结构是在目录树的帮助下确定的。解释性注释以增量方式处理。插图、注解、链接和文献引文也按优先级顺序以增量方式处理。增量写作是一个开放式过程,可在任何阶段中断,以便将文章导出为单一文档进行文本流程上的改写。用增量写作写成的文章可能特别适合增量阅读。它们可以与维基百科相比较。维基百科是增量阅读的极佳来源,因其增量增长和扎实的局部上下文。出于完全相同的原因,用增量写作汇编的材料也非常适合增量阅读。它们可能冗长且重复,然而借助增量阅读,可以以合理方式对它们进行优先级排序。增量写作使文本高度颗粒化,思路流向参差不齐,但在增量阅读中这是优点,因为所有独立文章和子文章都带有足够的局部上下文,可以独立阅读。更多内容见:Incremental writing。
致谢(Acknowledgements)
如果没有朋友、合作伙伴和家人的多方贡献,本文将不可能完成。我特别感谢 Miko Hejwosz、Dariusz Murakowski、Aleksandra Wozniak 博士、Edward Gorzelanczyk 博士、Janusz Murakowski 博士、Monika Morawska、Krzysztof Biedalak 和 Kuba Chodakowski 的帮助、耐心、贡献和想法。此外,无数个人通过提交他们的睡眠数据、回答我那些常常需要透露高度个人信息的问题,为我的工作提供了帮助。
术语表(Glossary)
本术语表采用的定义经过有意的简化或过度简化,以传递重要信息。如需更精确的定义,请跟随所提供的链接或自行检索:
acrophase(峰值相位)
正弦波的峰值。在本文中主要用于指”深夜”或”昼夜节律最低点”
adenosine(腺苷)
一种在清醒活动期间于大脑中积累、诱发睡意的物质(见:Adenosine)。其拮抗剂是咖啡因。
Ambien
流行的安眠药
apnea(呼吸暂停)
呼吸的缺失
提前睡眠相位综合征(advanced sleep phase syndrome),患者倾向于很早就寝并在日出前许久醒来的状况
脑源性神经营养因子(brain-derived neurotrophic factor),促进大脑生长的蛋白质(随睡眠、学习和运动而增加)
BF
Borbely(Borbély)
匈牙利睡眠科学家,以他的睡眠倾向双过程模型闻名
Buzsaki(Buzsáki)
匈牙利睡眠科学家,以他的REM 与 NREM 中反向信息流理论闻名
benzodiazepines(苯二氮䓬类)
一类流行的安眠药
biphasic sleep(双相睡眠)
由两个每日睡眠时段组成的睡眠。在本文中,双相睡眠被理解为一次长夜间睡眠加一次短白天小睡的组合。双相睡眠的其他变体也存在,但本文不使用其他含义(例如分段睡眠也可以是双相的)
caffeine(咖啡因)
咖啡、茶和可乐中的活性化合物,使人更警觉
catastrophic forgetting(灾难性遗忘)
人工神经网络中因干扰而可能发生的大规模遗忘。一种理论认为,人类在睡眠中使用双网络系统来防止灾难性遗忘。见:睡眠中的神经优化
chronotype(睡眠节律型/时型)
使人成为百灵鸟、猫头鹰、长睡眠者、不规律睡眠者等的睡眠控制系统特征
circadian sleep component(昼夜节律睡眠组分)
驱动人们入睡的两条主要睡眠倾向组分之一。昼夜节律组分通常在夜间表达,并决定夜间睡眠的时机
circadian cycle(昼夜节律周期) - 身体中约 24 小时的变化周期
circadian rhythm(昼夜节律) - 昼夜节律周期中变化的节律
circadian phase(昼夜节律相位) - 相对于昼夜节律周期的一天中的时间
circadian shift(昼夜节律相位移) - 昼夜节律周期相位的移动
circadian low(昼夜节律低谷) - 昼夜节律周期中我们感觉最困的时间
circadian curve(昼夜节律曲线) - 表示昼夜节律周期的正弦波(其高低代表困倦程度)。它可能是两道或更多正弦波的叠加,尤其是在年龄较小时
circadian preferences(昼夜节律偏好) - 对睡眠时机的偏好(例如百灵鸟喜欢早睡)
complementary encoding theory(互补编码理论)
一种记忆理论,认为安全编码记忆需要两套互补的记忆系统:快速、低干扰、小型临时系统,以及缓慢、重叠、庞大的长期系统。新记忆抵达快速系统,并高效地重新分配到庞大系统中。那种重新分配很可能发生在睡眠中。海马体和新皮层是互补系统的典型例子
consolidation(巩固)
-
记忆:在 SuperMemo 中,记忆通过复习得到强化的过程。在 SleepChart 中,可显示睡眠对记忆巩固的影响
-
睡眠时段:合并相邻的睡眠时段,以消除短暂夜间醒来对计算昼夜节律最低值过程的影响(在 SleepChart 中)
cry it out method(“哭到睡着”法)
一种通过让婴儿学到”为妈妈哭泣是徒劳的”来让婴儿入睡的方法。这种”方法”违背人类生物学,并可能对婴儿的大脑发育产生负面影响
DMH
下丘脑背内侧核(dorsal medial nucleus of hypothalamus),负责将昼夜节律信号与来自其他脑中心的信息整合以控制昼夜节律行为和昼夜节律生理的脑中心
DR
中缝背核(dorsal raphe),大脑的警觉中枢之一
延迟睡眠相位综合征(delayed sleep phase syndrome),每天醒得越来越晚的倾向
dual network theory(双网络理论)
一种睡眠理论,认为大脑发展出了一套双网络系统:一个用于短期低干扰记忆,另一个用于长期记忆存储。记忆向长期存储的转移发生在睡眠中
人的自然睡眠周期被调整以适应 24 小时日、且醒来时间足够早以满足工作、上学、社交等需要的过程。驯化的睡眠(entrained sleep)是不需要闹钟来”使其归位”的睡眠
farmer’s lifestyle(农夫式生活方式)
一种有利于好睡眠的生活方式。它包括早起(可能伴随日出)、大量户外体力劳动(或运动),以及一个没有电的放松夜晚(例如在壁炉的昏暗灯光下)。背离这种古老生活方式是睡眠障碍流行的根源。
free running sleep(自由睡眠)
不被人为控制(例如用安眠药或闹钟)的睡眠
garbage collection theory(垃圾回收理论)
一种认为睡眠的主要功能是清理记忆中信息垃圾的理论。这一理论在很大程度上是正确的。见:睡眠中的神经优化
habitual napper(习惯性小睡者)
把小睡作为常规习惯的人。习惯性小睡者不需要疲倦就能获得一次好小睡。在睡眠文献中,习惯性小睡者常被称为食欲性小睡者(appetitive nappers)
大脑的一小部分,对长期记忆的发展必不可少。它参与睡眠中的记忆优化。海马体受损的人可以睡眠,但他们会迅速忘记几乎所学的全部内容
又名食欲素(orexins),大脑中一种重要的警觉激素,参与调节睡眠。影响食欲素释放的基因突变可导致嗜睡症
homeostatic sleep component(内稳态睡眠组分)
睡眠倾向的组分。内稳态睡意可以比作”清醒的疲倦”。它在一天中逐渐增加
homeostatic process(内稳态过程) - 睡意在一天中随脑力消耗成比例增加的过程(文献中常记为 Process S)
homeostatic sleepiness(内稳态睡意) - 由脑力消耗导致的睡意(相对于通常在夜间到来的昼夜节律睡意)
homeostasis(稳态) - 身体响应外部刺激而控制其内部变量的过程(例如 REM 稳态是与 REM 睡眠亏缺成比例的 REM 睡眠驱动的积累)
hypnogram(睡眠图)
显示整夜睡眠阶段进展的图
insomnia(失眠)
入睡或维持睡眠的问题(见:Insomnia)
LC
蓝斑核(locus coeruleus),大脑的警觉中枢之一
melatonin(褪黑素)
夜间在大脑中释放的天然睡眠激素
monophasic sleep(单相睡眠)
24 小时内只包含一次长夜间睡眠时段的睡眠模式
MPO
亦作 MPA、MPOA
内侧视前区(medial preoptic area),负责整合睡眠启动所需信息的脑中心之一。MPOA 也参与育儿和性行为(包括性偏好)
NREM
非 REM 睡眠,通常开启夜间睡眠
orexins(食欲素)
orexin receptor(食欲素受体)
检测食欲素/下丘脑分泌素存在的受体
phase shift(相位移)
昼夜节律周期相位的变化。例如,1 小时的相位延迟等同于就寝时间推迟一小时、醒来时间推迟一小时、最低体温推迟一小时等。
pineal gland(松果体)
大脑中参与睡眠控制的一群细胞。它负责夜间褪黑素的释放
polyphasic sleep(多相睡眠)
由一天内多次睡眠时段组成的睡眠模式(在本文中,双相睡眠不算作多相睡眠模式的一部分)
PRC
相位响应曲线(phase response curve),一条说明身体如何响应使睡眠相位移动的信号的曲线(例如傍晚的光照如何让我们更晚犯困、更晚醒来)
PVN
亦作 PVH
下丘脑室旁核(paraventricular nucleus of the hypothalamus),负责整合睡眠启动刺激的脑中心之一
RAS
网状激活系统(reticular activating system,位于脑干),负责整体唤起的大脑部分
recall(回忆)
从记忆中调出信息的能力(见:Memory recall)
REM
快速眼动(rapid eye movement)睡眠
REM deficit(REM 亏缺)
所获 REM 睡眠量的欠债。这笔债必须尽快偿还,但通常在 NREM 睡眠亏缺偿还之后
reverse learning theory(反向学习理论)
一种认为我们在睡眠中遗忘较不重要事物的理论(见:Sleep as neural optimizer)
siesta(午睡)
正午的小睡
sleep apnea(睡眠呼吸暂停)
睡眠中呼吸的缺失
用于绘制睡眠的免费应用程序
sleep deprivation(睡眠剥夺)
慢性或急性的睡眠不足
sleep latency(睡眠潜伏期)
上床与入睡之间的时间
SPW
在深睡中出现、可能与短期记忆向新皮层转移相关的大脑波(sharp-wave,尖锐波)
SWS
慢波睡眠(slow-wave sleep),睡眠的最深阶段,对新经验与旧记忆的巩固必不可少。NREM 睡眠的第 3-4 阶段
subjective night(主观夜间)
身体想要睡眠的时间。它通常发生在夜间,但对猫头鹰而言同样可能是清晨、甚至正午
使每个人都能研究自己睡眠与学习之间关系的加速学习程序
THC
四氢大麻酚(tetrahydrocannabinol),大麻中发现的精神活性化合物
TMN
下丘脑结节乳头核(tuberomamillary nucleus),大脑的警觉中枢之一
TMS
经颅磁刺激(transcranial magnetic stimulation),一种借助磁场无需手术即可刺激大脑部分的方法
腹外侧视前核(ventrolateral preoptic nucleus),负责启动睡眠的主要中枢之一
总结(Summary)
睡眠的 ABC(ABC of sleep)
- 睡眠对学习、创造力至关重要! 睡眠剥夺导致智力剥夺!
- 想睡多少睡多少。在对齐良好的昼夜节律周期中,你不可能睡得过多。
- 睡眠不是食物,你不能提前获得更多并储存以备后用(见:Excessive sleeping)。
- 避免闹钟,它们对学习和健康有害!
- 忘掉试图在预定时间入睡!让身体决定!
- 忘掉试图快速入睡!如果身体决定是正确时间,快速睡眠会自然到来!
- 不要试图让自己困倦!只要保持清醒并持续工作/学习足够久即可,自然睡眠会到来!(见:Insomnia)
- 消除压力源比试图在睡前忘记压力情境要好得多!(见:Stress)
- 如果你 30 分钟无法入睡,起床!你还没准备好睡眠!(见:Insomnia)
- 记住,糟糕的一夜是生活的因素,很少有人能避免。然而,一丝不苟地遵守睡眠卫生规则将帮助你大幅减少糟糕一夜的发生。
- 如果你坚持健康睡眠规则,你应不知道什么是睡眠惰性。
- 即便轻微程度的睡眠剥夺也会在数天内累积,严重降低表现。
- 睡眠剥夺给社会带来天文数字的成本。
- 轮班工作和时差对健康有害!
- 成年人是双相的,但婴儿是多相的。不小睡者也在中午显示双相个体典型的学习表现低谷。
优化睡眠(Optimizing sleep)
- 对抗失眠的最佳武器是恰当的睡眠时机,你应只在足够疲倦时入睡。
- 如果你想最大化处于高效清醒状态的时间,请阅读关于自由睡眠的内容,让你的生物学为你工作,而非与你对抗。
- 你不应最小化睡眠,而应优化睡眠量,这意味着你应恰好获得履行其神经功能所需的睡眠。
- 了解你自己睡眠时机的细节:睡多少小时、需要小睡前多少小时或再次入睡等。用本文找出如何计算你的最佳作息。用 SleepChart 获得困难情况的可视化辅助,用 SuperMemo 查看睡眠如何影响你的学习。
- 每个人在中午(大约自然醒来后 7 小时)都经历学习能力的大幅低谷,这是午睡的好时机。不要在自然醒来 8 小时之后小睡。
- 如果你睡够了仍不感觉神清气爽,确保你没有违背昼夜节律睡眠,尝试自由睡眠。记住,你可能需要 1-2 周来同步所有身体功能才能完美工作!
- 如果在尝试至少一个月自由睡眠后仍无法获得令人神清气爽的睡眠,咨询睡眠专家。记住,即便自由睡眠也可能对有睡眠相位障碍或高度不规律睡眠的人有挑战,然而你大概有 90-95% 的机会它将回答你大部分的睡眠质量问题!
- 即便自由睡眠几乎是好睡眠的近乎保证,自由运行作息可能与你的工作或社交生活不兼容。
- 没有睡眠剥夺,你自由运行时最小化总睡眠。人类过度睡眠的倾向是迷思。(见:Excessive sleeping)
睡眠与学习(Sleep and learning)
- 闹钟对学习有害!
- 你的学习能力取决于你的生物钟,而非墙上挂的时钟。
- 你应根据昼夜节律周期调整智力工作的时机。
- 睡眠中学习的磁带是营销噱头。
- 健康时机恰当的睡眠后,最佳回忆发生在清晨时段。如果你是小睡者,第二个良好学习时段在午睡之后。(见:Recall)
- 健康时机恰当的睡眠后,最佳记忆巩固(最佳学习)发生在清晨时段。(见:Memory consolidation)
- 为获得最佳学习结果,你需要在学习前后都有良好睡眠(Walker,2008122)。
- 繁重学习阻碍学习本身,使填鸭式复习双重适得其反:(1) 填鸭留下很少长期记忆痕迹;(2) 填鸭快速抑制短期记忆。
- 仅仅警觉和敏锐警觉之间的学习能力有巨大差异。
- 中午学习能力有大幅低谷,那时小睡比学习更好!(见:Biphasic nature of human sleep)
- 小睡对学习极好。不要把双相睡眠的自然小睡与”设计师小睡”(如多相”Uberman”睡眠)混淆!
- 放松状态下的学习工具 conducive 小睡,它们对实际学习的有用性有限。
一般(General)
- 睡眠对大脑生长极好。
- 如果选择恰当、时机恰当,运动对睡眠有益,睡眠也对运动有益。
- 学习对睡眠有益,睡眠对学习有益。
- 压力、酒精、娱乐性毒品和安眠药都对睡眠有害,尤其是在睡前不久使用时。
- 与好人相处!很少有东西能像坏人一样有效地毁掉睡眠!
- 如果你在身体呼唤睡眠时经历思绪飞驰,最佳方法是:起床做些在智力上让你疲倦的事!
- 如果你想为睡眠研究做贡献,或想更多了解自己的睡眠和学习,或有该主题的新想法,请写信给我。
- 并非所有科学家都同意本文呈现的事实(见:Jim Horne 和 Daniel Kripke、Robert Vertes 和 Jerome Siegel)。
儿童(Children)
- 让婴儿按需睡眠,共同睡眠对他们的情感和智力发育至关重要。
- 给定基本安全措施,同床共眠对婴儿极好,并帮助发展他们的昼夜节律模式(只要妈妈自己不违反睡眠卫生规则)。
- 永远不要叫醒婴儿或小孩!这对他们的大脑生长有害!除非儿科医生特别指示,喂养作息必须让位。睡眠模式应决定婴儿一天的组织方式。
- 如果你必须送孩子去幼儿园,确保驯化适当的睡眠模式,永远不需要叫醒孩子!这对他们的大脑生长至关重要!如果你无法驯化他们的睡眠,考虑其他选择(包括在家护理孩子)。
小睡(Napping)
- 小睡改善脑力表现。
- 一天一次小睡足够。
- 小睡应在一天的第 7 小时进行,较早的小睡低效,较晚的小睡可能搞乱昼夜节律周期(见:Best nap timing)。
- 小睡无法弥补夜间睡眠剥夺,然而它们对休息良好的大脑是绝佳提升!(见:Optimum nap duration)
物质(Substances)
- 小心咖啡因,只在醒来时(或小睡后,如果你小睡)喝咖啡。
- 每天不要超过一杯酒(除非你大量运动),避免睡前饮酒(尤其是在夜间睡眠前最后 2-3 小时)。
- 戒烟!
- 大麻是亦敌亦友!它可能帮助你入睡,但那种睡眠不会令人神清气爽!
- 褪黑素可以帮助你更早入睡并前移睡眠相位,但它也会导致不太令人神清气爽的睡眠。
睡眠的作用(Role of sleep)
- 睡眠的主要功能是优化存储在大脑神经网络中的记忆。
- 如果可强制执行,睡眠剥夺会导致死亡(可能在几周内)。
- 睡眠剥夺的致命结果可能由主要的神经网络故障引起,也可能由睡眠保护程序的次级效应造成(见:Why do we die without sleep?)。
- 从不睡眠的健康人是迷思,要么他们睡要么他们死。
DSPS
- 年轻求学一代中相当一部分受某种程度的 DSPS 障碍影响。
- DSPS 病例的流行病增加最可能由基于使用电力的现代生活方式引起,它破坏了传统的光暗周期及相关社会线索(见:Is DSPS a disease?)。
- 正常人可能在自由睡眠作息上经历相位移,揭示了现代生活方式与睡眠生物学之间流行的错配。
多相睡眠(Polyphasic sleep)
- 多相睡眠导致混乱的相位移响应。
- 在多相睡眠尝试中,昼夜节律周期很可能像在无授时因子条件下一样在后台运行,混乱的相位移输入互相抵消。
- 健康人无法驯化多相睡眠,也无法在不睡眠剥夺的情况下维持多相作息。不可能多相睡眠并同时长期保持最大创造力、警觉性和健康。
- 任何声称处于永久多相作息的人要么患有睡眠障碍,要么不诚实。
睡眠生理学(Physiology of sleep)
- 我们只有在两个条件满足时才进入健康睡眠:(1) 清醒带来的疲倦高(高内稳态睡眠倾向),以及 (2) 生物钟说”该睡觉了”(高昼夜节律睡眠倾向)(见:Borbély model)。
- 睡眠像双稳态开关一样运作,具有快速过渡和稳定的睡眠或清醒阶段。当睡眠卫生规则被违反时,这些健康的快速过渡可能效率较低。
- 睡眠在夜间在两种不同模式间交替:NREM 睡眠和 REM 睡眠,它们以约 90 分钟周期不断变化。两个阶段都非常重要,在学习和创造力过程中扮演不同角色。
- 大脑在睡眠中经历重大重组,一些突触权重被减弱,另一些被强化,神经联结被优化为具有最小干扰的一致抽象关联记忆。
- 人类睡眠可以通过时间疗法相对于时钟重新定位,睡眠相位的变化沿相位响应曲线(PRC)进行。
来源(Sources)
本文主要从 supermemo.com 的其他来源汇编而成,包括:
- Roots of creativity and genius
- FAQ: Good sleep for good learning
- SleepChart: Formula for healthy sleep
- Polyphasic sleep: myths and facts
- Good sleep, good learning, good life (2000)
参考文献精选(Selected references)
以下是本系列引用的部分关键参考文献(完整参考文献列表见原文):
- Stickgold R., Sleep-dependent memory consolidation, Nature 437 (2005): 1272-1278.
- Buzsáki G., Two-stage model of memory trace formation, Neuroscience 31(3) (1989): 551-570.
- Borbély A.A., A two process model of sleep regulation, Human Neurobiology 1(3) (1982): 195-204.
- Rechtschaffen A., Current perspectives on the function of sleep, Perspectives in Biology and Medicine 41(3) (1998): 359-390.
- Siegel J.M., Why we sleep, Scientific American 289(5) (2003): 92–97.
- Saper C.B., Scammell T.E., Lu J., Hypothalamic regulation of sleep and circadian rhythms, Nature 437 (2005): 1257-1263.
- Czeisler C.A. 等, Stability, Precision, and Near-24-Hour Period of the Human Circadian Pacemaker, Science 284(5423) (1999): 2177-2181.
- Wehr T.A., In short photoperiods, human sleep is biphasic (1992).
- Dinges D.F., Broughton R.J., Sleep and Alertness: Chronobiological, Behavioral, and Medical Aspects of Napping (纽约:Raven Press,1989).
- Stampi C., Why We Nap: Evolution, Chronobiology and Functions of Polyphasic and Ultrashort Sleep (波士顿:Birkhäuser,1992).
- Knutson K.L. 等, The metabolic consequences of sleep deprivation, Sleep Medicine Reviews 11(3) (2007): 163-178.
- Walker M.P., Stickgold R., It’s Practice, with Sleep, that Makes Perfect, Clinics in Sports Medicine 24(2) (2005): 301-317.
- Tononi G., Cirelli C., Sleep function and synaptic homeostasis, Sleep Medicine Reviews 10(1) (2006): 49-62.
- McClelland J.L., McNaughton B.L., O’Reilly R.C., Why there are complementary learning systems in the hippocampus and neocortex, Psychological Review 102(3) (1995): 419-457.
- Wilson M.A., McNaughton B.L., Reactivation of hippocampal ensemble memories during sleep, Science 265(5172) (1994): 676-679.
- Wagner U. 等, Sleep inspires insight, Nature 427 (2004): 352-355.
- Vertes R.P., Siegel J.M., Time for the Sleep Community to Take a Critical Look at the Purported Role of Sleep in Memory Processing, Sleep 28(10) (2005): 1228-1229.
- Cappuccio F.P. 等, Sleep Duration and All-Cause Mortality, Sleep 33(4) (2010): 585-592.
- Hobson J.A., McCarley R.W., The brain as a dream state generator, Am. J. Psychiatry 134(12) (1977): 1335-1348.
- Crick F., Mitchison G., The function of dream sleep, Nature 304 (1983): 111-114.
译注:本系列为 Dr Piotr Wozniak《Good sleep, good learning, good life》的忠实中译,原文见 supermemo.guru。译文保留作者第一人称视角与论证脉络,关键技术术语保留英文原名以便对照。如对译文有任何疑问或建议,欢迎反馈。
-
国家睡眠障碍研究委员会(National Commission on Sleep Disorders Research),Wake Up America: A National Sleep Alert,美国卫生与公众服务部出版物,1992。指出睡眠剥夺每年因更高的压力和更低的工作场所生产力而造成约 1500 亿美元损失。 ↩
-
Mitler M.M., Carskadon M.A., Czeisler C.A., Dement W.C., Dinges D.F., Graeber R.C., Catastrophes, Sleep, and Public Policy: Consensus Report,Sleep,第 11 卷第 1 期(1988):100–109。指出包括三哩岛、切尔诺贝利、博帕尔、Zeebrugge、Exxon Valdez 在内的重大工业灾难均与相关人员睡眠剥夺直接相关。 ↩
-
Stickgold R., Sleep-dependent memory consolidation,Nature,第 437 卷(2005 年 10 月 27 日):1272-1278。证明睡眠对学习是必需的。 ↩ ↩2
-
Buzsáki G., Two-stage model of memory trace formation: A role for “noisy” brain states,Neuroscience,第 31 卷第 3 期(1989):551-570。关于睡眠中记忆痕迹形成的两阶段模型。 ↩ ↩2
-
Wozniak P.A., Gorzelanczyk E.J., Murakowski J.A., Two components of long term memory,Acta Neurobiologiae Experimentalis,第 55 卷(1995):301-305。 ↩
-
Borbély A.A., A two process model of sleep regulation,Human Neurobiology,第 1 卷第 3 期(1982):195-204。睡眠调节的双过程模型。 ↩ ↩2
-
Rechtschaffen A., Current perspectives on the function of sleep,Perspectives in Biology and Medicine,第 41 卷第 3 期(1998):359-390。记录了完全剥夺睡眠的大鼠平均在 2.5 周内死亡(仅剥夺 REM 睡眠约 5 个月)。 ↩
-
Siegel J.M., Why we sleep,Scientific American,第 289 卷第 5 期(2003):92–97。证明睡眠剥夺大鼠存在脑损伤。 ↩
-
Hairston I.S., Little M.T.M., Scanlon M.D., Barakat M.T., Palmer T.D., Sapolsky R.M., Heller H.C., Sleep restriction suppresses neurogenesis induced by hippocampus-dependent learning,Journal of Neurophysiology,第 94 卷第 6 期(2005 年 12 月):4224-4233。指出睡眠不足抑制大脑某些区域的神经发生。 ↩
-
Frank M.G., Issa N.P., Stryker M.P., Sleep Enhances Plasticity in the Developing Visual Cortex,Neuron,第 30 卷第 1 期(2001 年 4 月):275-287。 ↩ ↩2 ↩3
-
Whitfield J., To sleep, perchance to learn,Nature(2001 年 4 月),doi:10.1038/news010426-15。引述 Stryker 博士关于夜间激素变化在巩固和增强清醒经验中的作用。 ↩
-
Rhodes & Associates Inc., Fatigue Management Guide for Canadian Marine Pilots – A Trainer’s Handbook(Wayne Rhodes,2001)。调查显示只有 10% 的轮班工作者对其睡眠作息没有抱怨。 ↩
-
Cho K., Chronic ‘jet lag’ produces temporal lobe atrophy and spatial cognitive deficits,Nature Neuroscience,第 4 卷第 6 期(2001 年 6 月):567-568。长期洲际飞行的空乘人员海马体出现细胞丢失。 ↩
-
Cho K., Ennaceur A., Cole J.C., Suh C.K., Chronic Jet Lag Produces Cognitive Deficits,The Journal of Neuroscience,第 20 卷第 6 期(2000 年 3 月 15 日):RC66。 ↩
-
Knutson K.L., Spiegel K., Penev P., Van Cauter E., The Metabolic Consequences of Sleep Deprivation,Sleep Medicine Reviews,第 11 卷第 3 期(2007 年 6 月):163-178。 ↩
-
Allan J.S., Czeisler C.A., Persistence of the circadian thyrotropin rhythm under constant conditions and after light-induced shifts of circadian phase,The Journal of Clinical Endocrinology & Metabolism,第 79 卷第 2 期(1994 年 8 月 1 日):508-512。 ↩
-
Baumgartner A. 等,Influence of partial sleep deprivation on the secretion of thyrotropin, thyroid hormones, growth hormone, prolactin, luteinizing hormone, follicle stimulating hormone, and estradiol in healthy young women,Psychiatry Research,第 48 卷第 2 期(1993 年 8 月):153–178。 ↩
-
Mills E., Wu P., Seely D., Guyatt G., Melatonin in the treatment of cancer: a systematic review of randomized controlled trials and meta-analysis,Journal of Pineal Research,第 39 卷第 4 期(2005 年 11 月):360-366。 ↩
-
Schernhammer E.S. 等,Epidemiology of urinary melatonin in women and its relation to other hormones and night work,Cancer Epidemiology, Biomarkers & Prevention,第 13 卷第 6 期(2004 年 6 月):936-943。 ↩
-
Dijk D.J., Lockley S.W., Integration of human sleep-wake regulation and circadian rhythmicity,Journal of Applied Physiology,第 92 卷第 2 期(2002):852-862。指出要获得超过 8 小时不间断睡眠,应在体温最低点前约 6 小时启动睡眠。 ↩
-
Wozniak P.A., Gorzelanczyk E.J., Formula for healthy sleep(2003)。介绍 SleepChart 工具用于可视化绘制睡眠。 ↩
-
Roberts R.D., Kyllonen P.C., Morningness–eveningness and intelligence: early to bed, early to rise will likely make you anything but wise!,Personality and Individual Differences,第 27 卷第 6 期(1999 年 12 月):1123-1133。研究表明猫头鹰比百灵鸟表现出略高的智商。 ↩
-
Dinges D.F., Broughton R.J., Sleep and Alertness: Chronobiological, Behavioral, and Medical Aspects of Napping(纽约:Raven Press,1989),171-204。 ↩ ↩2
-
Dinges D.F., “Adult napping and its effects on ability to function.” 载于 C. A. Stampi 编《Why we Nap》(波士顿:Birkhäuser,1992),118–134。 ↩
-
Wertz A.T., Ronda J.M., Czeisler C.A., Wright K.P. Jr, Effects of Sleep Inertia on Cognition,JAMA,第 295 卷第 2 期(2006):163-164。 ↩
-
Wehr T.A., In short photoperiods, human sleep is biphasic(1992)。Thomas Wehr 关于延长黑暗时段分段睡眠的经典实验。 ↩ ↩2
-
Wehr T.A. 等,Conservation of photoperiod-responsive mechanisms in humans,American Journal of Physiology,第 265 卷第 4 期(1993 年 10 月):846-857。 ↩
-
Ekirch A.R., Sleep We Have Lost: Pre-industrial Slumber in the British Isles,The American Historical Review,第 106 卷第 2 期(2001):343-387。指出电力之前的历史记录频繁提及两段式睡眠。 ↩
-
Carskadon M.A., “Early school schedules modify adolescent sleepiness,” Sleep Research,第 24 卷:92。指出 DSPS 在青春期非常普遍。 ↩ ↩2
-
Golombek D.A., Rosenstein R.E., Physiology of Circadian Entrainment,Physiological Reviews,第 90 卷第 3 期:1063-1102。影响时钟周期的突变。 ↩ ↩2 ↩3 ↩4
-
Neubauer D.N., “Delayed sleep phase syndrome: Analysis of a 10-year sleep log,” Johns Hopkins Sleep Disorders Center(2000)。记录有人沿移动 DSPS 作息生活数十年而无重大健康副作用。 ↩
-
LivingWithN24, Charting the course of N24,DSPS, a sleep disorder(2010 年 10 月 27 日)。同时表明不规律的”自由”睡眠会使睡眠模式越来越不规律。 ↩ ↩2
-
Wyatt J.K., Ritz-De Cecco A., Czeisler C.A., Dijk D.J., Circadian temperature and melatonin rhythms, sleep, and neurobehavioral function in humans living on a 20-h day,American Journal of Physiology,第 277 卷第 4 期(1999 年 10 月):1152-1163。20 小时日强制失步协议。 ↩
-
Carskadon M.A., Labyak S.E., Acebo C., Seifer R., Intrinsic circadian period of adolescent humans measured in conditions of forced desynchrony,Neuroscience Letters,第 260 卷第 2 期(1999 年 1 月 29 日):129–132。28 小时日强制失步协议。 ↩
-
Weitzman E.D. 等,Delayed sleep phase syndrome. A chronobiological disorder with sleep-onset insomnia,Archives of General Psychiatry,第 38 卷第 7 期(1981 年 7 月):737–746。假设相当数量的入睡性失眠患者可能患有未诊断的 DSPS。 ↩
-
Sensi S., Capani F., Chronobiological Aspects of Weight Loss in Obesity: Effects of Different Meal Timing Regimens,Chronobiology International,第 4 卷第 2 期(1987):251-261。 ↩
-
Rehm D.S., The Myopia Myth(Ligonier:International Myopia Prevention Association,1981)。支持近视欠矫正的理论。 ↩
-
Coghlan A., Le Page M., Eye correction is seriously short sighted,New Scientist(2002 年 11 月 20 日)。提供与欠矫正相反的证据。 ↩
-
Sack R. 等,Circadian rhythm sleep disorders: Part II,SLEEP,第 30 卷第 11 期(2007):1484-1501。 ↩ ↩2
-
Hayakawa T. 等,Trials of bright light exposure and melatonin administration in a patient with non-24 hour sleep-wake syndrome,Psychiatry and Clinical Neurosciences,第 52 卷第 2 期:261–262。指出无法维持 24 小时睡眠-清醒周期的人中 25% 患有精神障碍。 ↩
-
Kripke D.F. 等,Delayed sleep phase cases and controls,Journal of Circadian Rhythms,第 6 卷第 6 期(2008)。得出 DSPS 表型家族性并与单相抑郁症相关。 ↩
-
Rao M.R. 等,Long term cognitive development in children with prolonged crying,Archives of Disease in Childhood,第 89 卷第 11 期(2004):989-992。长时间哭泣与后来的认知缺陷相关。 ↩
-
Odaa G.A. 等,Breastfeeding, sleep and wake circadian rhythms show distinct temporal emerging patterns,Biological Rhythm Research,第 39 卷第 4 期(2008):379-387。 ↩
-
Rivkees S.A., Fox C.A., Jacobson C.D., Reppert S.M., Anatomic and Functional Development of the Suprachiasmatic Nuclei in the Gray Short-Tailed Opossum,The Journal of Neuroscience,第 8 卷第 11 期(1988 年 11 月):4289-4276。 ↩ ↩2
-
Cubero J. 等,The circadian rhythm of tryptophan in breast milk affects the rhythms of 6-sulfatoxymelatonin and sleep in newborn,Neuroendocrinology Letters,第 26 卷第 6 期(2005):657-661。母乳中色氨酸的昼夜节律影响婴儿褪黑素与睡眠。 ↩
-
Altman J., Bayer S.A., Development of the diencephalon in the rat. I.,The Journal of Comparative Neurology,第 182 卷第 5 期(1978 年 12 月 15 日):945-997。大鼠 SCN 周期在子宫内发育。 ↩
-
Page T.L., Developmental manipulation of the circadian pacemaker in the cockroach,Physiological Entomology,第 16 卷第 2 期(1991 年 6 月):243-248。相位响应曲线形状受发育期间照明条件影响。 ↩
-
McKenna J.J., McDad T., Why babies should never sleep alone,Paediatric Respiratory Reviews,第 6 卷第 2 期(2005 年 6 月):134-152。推荐母婴共同睡眠。 ↩
-
McKenna J.J., Is sleeping with my baby safe?,Horizons,第 1 卷第 4 期(1995 年春/夏)。 ↩
-
Walker M.P., Stickgold R., It’s Practice, with Sleep, that Makes Perfect,Clinics in Sports Medicine,第 24 卷第 2 期(2005 年 4 月):301-317。 ↩
-
Nishida M., Walker M.P., Daytime Naps, Motor Memory Consolidation and Regionally Specific Sleep Spindles,PLoS ONE,第 2 卷第 4 期(2007):e341。 ↩
-
Werth E., Dijk D.J., Achermann P., Borbély A.A., Dynamics of the sleep EEG after an early evening nap,American Journal of Physiology,第 271 卷第 3 期(1996 年 9 月):501-510。 ↩
-
Stampi C., Why We Nap: Evolution, Chronobiology and Functions of Polyphasic and Ultrashort Sleep(波士顿:Birkhäuser,1992)。指出多相作息最难的部分是从昼夜节律顶点处的小睡中醒来。 ↩ ↩2
-
Czeisler C.A. 等,Stability, Precision, and Near-24-Hour Period of the Human Circadian Pacemaker,Science,第 284 卷第 5423 期(1999 年 6 月 25 日):2177-2181。测量人类昼夜节律周期为 24.2 小时,个体间变化惊人地小。 ↩
-
Khalsa S.B.S. 等,A phase response curve to single bright light pulses in human subjects,The Journal of Physiology,第 549 卷第 3 期(2003 年 6 月 15 日):945-952。 ↩
-
Ruby N.F. 等,Siberian hamsters free run or become arrhythmic after a phase delay of the photocycle,American Journal of Physiology,第 271 卷第 4 期(1996 年 10 月):881-890。通过光刺激使西伯利亚仓鼠无节律。 ↩ ↩2 ↩3
-
Thaler J.P. 等,Obesity is associated with hypothalamic injury in rodents and humans,The Journal of Clinical Investigation,第 122 卷第 1 期(2012 年 1 月 3 日):153–162。指出垃圾饮食如何对控制食欲的脑中心造成胶质损伤。 ↩
-
Swaab D.F. 等,Development of vasopressin neurons in the human suprachiasmatic nucleus in relation to birth,Brain research. Developmental brain research,第 52 卷(1990):289-293。新生儿 SCN 只含成年血管加压素表达神经元数量的 13%。 ↩
-
Swaab D.F. 等,The human suprachiasmatic nucleus (SCN) in a periodic environment,Proceedings of the fifth Sapporo Symposium of Biological Rhythm(1994):323-337。 ↩ ↩2
-
Mirmiran M., Ariagno R.L., Influence of light in the NICU on the development of circadian sleep component rhythms in preterm infants,Seminars in Perinatology,第 24 卷第 4 期(2000 年 8 月):247-257。 ↩
-
Wehr T.A.,1992。分段睡眠实验,谈论的是非常长的睡眠块,而非 Uberman 式的迷你小睡。 ↩
-
Wozniak P.A., Polyphasic Sleep: Facts and Myths(2005)。 ↩
-
Zimmermann T., Miles to go before I sleep,Outside Magazine(2005 年 4 月)。引述 Stampi 博士称”我从不疲倦”。 ↩
-
Pryor M., MacArthur is caught not napping,The Times(2005 年 1 月 20 日)。引述 Stampi 博士关于 Ellen MacArthur 的话。 ↩
-
Sbragia G., “Leonardo da Vinci and Ultrashort Sleep: Personal Experience of an Eclectic Artist.” 载于 C. A. Stampi 编《Why we Nap》(波士顿:Birkhäuser,1992),180–185。关于达芬奇睡眠习惯可能源自一位灵媒的故事。 ↩
-
Ruitenberg A. 等,Alcohol consumption and risk of dementia: the Rotterdam Study,The Lancet,第 359 卷第 9303 期(2002 年 1 月 26 日):281-286。适度饮酒可能降低阿尔茨海默病发病率。 ↩
-
Solinas M. 等,Caffeine Induces Dopamine and Glutamate Release in the Shell of the Nucleus Accumbens,The Journal of Neuroscience,第 22 卷第 15 期(2002 年 8 月 1 日):6321-6324。咖啡因影响奖励中心(包括伏隔核)。 ↩
-
Landolt H.P. 等,Caffeine intake (200 mg) in the morning affects human sleep and EEG power spectra at night,Brain Research,第 675 卷第 1-2 期(1995 年 3 月 27 日):67–74。即便早晨摄入咖啡因也会减少夜间深睡眠。 ↩
-
Van Praag H., Kempermann G., Gage F.H., Running increases cell proliferation and neurogenesis in the adult mouse dentate gyrus,Nature Neuroscience,第 2 卷第 3 期:266-270。 ↩
-
Guzmán-Marín R. 等,Sleep deprivation reduces proliferation of cells in the dentate gyrus of the hippocampus in rats,The Journal of Physiology,第 549 卷第 2 期(2003 年 6 月 1 日):563-571。 ↩
-
Gambelunghe C. 等,Effects of light physical exercise on sleep regulation in rats,Medicine & Science in Sports & Exercise,第 33 卷第 1 期(2001):57-60。 ↩
-
Feinberg I. 等,Effects of high dosage delta-9-tetrahydrocannabinol on sleep patterns in man,Clinical Pharmacology & Therapeutics,第 17 卷第 4 期(1975 年 4 月):458-466。 ↩
-
Fujimori M., Himwich H.E., delta9-Tetrahydrocannabinol and the sleep-wakefulness cycle in rabbits,Physiology & Behavior,第 11 卷第 3 期(1973 年 9 月):291-295。 ↩
-
Knutson K.L., Spiegel K., Penev P., Van Cauter E., The metabolic consequences of sleep deprivation,Sleep Medicine Reviews,第 11 卷第 3 期(2007 年 6 月):163-178。 ↩
-
Keim N.L. 等,Weight Loss is Greater with Consumption of Large Morning Meals,The Journal of Nutrition,第 127 卷第 1 期(1997 年 1 月 1 日):75-82。 ↩
-
Wren A.M. 等,The Novel Hypothalamic Peptide Ghrelin Stimulates Food Intake and Growth Hormone Secretion,Endocrinology,第 141 卷第 11 期(2000 年 11 月 1 日):4325-4328。禁食促进饥饿素释放。 ↩
-
Norrelund H., The metabolic role of growth hormone in humans with particular reference to fasting,Growth Hormone & IGF Research,第 15 卷第 2 期(2005 年 4 月):95-122。 ↩
-
Roberts S., Self-experimentation as a source of new ideas,Behavioral and Brain Sciences,第 27 卷第 2 期(2004 年 4 月):227-262。 ↩
-
Wozniak P.A., Can too much learning lead to Alzheimer’s?(2002 年 5 月)。 ↩ ↩2
-
SuperMemo World, SuperMemo User Survey(1994 年 11 月)。 ↩
-
Gorzelanczyk E.J., Wozniak P.A., Chrzastowska H., Investigating the correlation between the intelligence and the performance in repetitive learning tasks,第 3 届欧洲神经学会联合会大会,塞维利亚(1998)。 ↩
-
Cappuccio F.P. 等,Sleep Duration and All-Cause Mortality: A Systematic Review and Meta-Analysis,Sleep,第 33 卷第 4 期(2010):585-592。 ↩
-
Cappuccio F.P. 等,Quantity and Quality of Sleep and Incidence of Type 2 Diabetes,Diabetes Care,第 33 卷第 2 期(2010 年 2 月):414-420。 ↩
-
Wehr T.A.,1992。Wehr 的分段睡眠实验。 ↩
-
Dinges D.F., Rogers N.L., Baynard M.D., “Chronic Sleep Deprivation.” 载于 Kryger M.H. 等编《Principles and Practice of Sleep Medicine》(圣路易斯:Saunders,2000)。 ↩
-
De Koninck J. 等,Intensive language learning and increases in rapid eye movement sleep,International Journal of Psychophysiology,第 8 卷第 1 期(1989 年 9 月):43-47。 ↩
-
Smith C.T., Nixon M.R., Nader R.S., Posttraining increases in REM sleep intensity implicate REM sleep in memory processing,Learning Memory,第 11 卷(2004):714-719。 ↩ ↩2 ↩3
-
Wahistrom K., Changing Times: Findings From the First Longitudinal Study of Later High School Start Times,NASSP Bulletin,第 86 卷第 633 期(2002 年 12 月):3-21。 ↩
-
Hobson J.A., Pace-Schott E.F., Stickgold R., Dreaming and the brain: Toward a cognitive neuroscience of conscious states,Behavioral and Brain Sciences,第 23 卷第 6 期(2000):793-842。AIM 3D 睡眠-清醒空间模型。 ↩ ↩2
-
Wozniak P.A., The roots of creativity and genius(2001)。 ↩
-
Porkka-Heiskanen T., Adenosine in sleep and wakefulness,Annals of Medicine,第 31 卷第 2 期(1999):125-129。腺苷在基底前脑累积产生内稳态睡眠倾向。 ↩
-
Kong J. 等,Brain Glycogen Decreases with Increased Periods of Wakefulness,The Journal of Neuroscience,第 22 卷第 13 期(2002 年 7 月 1 日):5581-5587。 ↩
-
Scammell T.E. 等,An adenosine A2a agonist increases sleep and induces Fos in ventrolateral preoptic neurons,Neuroscience,第 107 卷第 4 期(2001 年 11 月 28 日):653–663。 ↩ ↩2 ↩3 ↩4
-
Krueger J.M., Obál F. Jr, Fang J., Humoral regulation of physiological sleep: cytokines and GHRH,Journal of Sleep Research,第 8 卷第 S1 期(1999 年 3 月):54-59。 ↩
-
Krueger J.M., Obál F. Jr, Fang J., Kubota T., Taishi P., The Role of Cytokines in Physiological Sleep Regulation,Annals of the New York Academy of Sciences,第 933 卷(2001 年 3 月):211-221。 ↩
-
De A., Krueger J.M., Simasko S.M., Glutamate induces the expression and release of tumor necrosis factor-alpha in cultured hypothalamic cells,Brain Research,第 1053 卷第 1-2 期(2005 年 8 月 16 日):54-61。 ↩
-
Daan S., Beersma D.G., Borbely A.A., Timing of human sleep: recovery process gated by a circadian pacemaker,American Journal of Physiology,第 246 卷第 2 期(1984 年 2 月):161-183。慢波活动作为内稳态睡眠倾向的标志。 ↩
-
Akerstedt T., Folkard S., “A model of human sleepiness.” 载于 J. A. Horne 编《Sleep ‘90》(Bochum:Pontenagel Press,1990),310-313。三过程模型基础。 ↩
-
Saper C.B., Chou T.C., Scammell T.E., The sleep switch: hypothalamic control of sleep and wakefulness,Trends in Neurosciences,第 24 卷第 12 期(2001 年 12 月 1 日):726-731。睡眠-清醒双稳态开关。 ↩
-
Saper C.B., Scammell T.E., Lu J., Hypothalamic regulation of sleep and circadian rhythms,Nature,第 437 卷(2005 年 10 月 27 日):1257-1263。 ↩
-
Swaab D.F., Hofman M.A., An enlarged suprachiasmatic nucleus in homosexual men,Brain Research,第 537 卷第 1-2 期(1990 年 12 月 24 日):141-148。 ↩
-
Chou T.C. 等,Critical Role of Dorsomedial Hypothalamic Nucleus in a Wide Range of Behavioral Circadian Rhythms,The Journal of Neuroscience,第 23 卷第 33 期(2003 年 11 月 19 日):10691-10702。 ↩
-
Gooley J.J., Schomer A., Saper C.B., The dorsomedial hypothalamic nucleus is critical for the expression of food-entrainable circadian rhythms,Nature Neuroscience,第 9 卷(2006):398-407。 ↩
-
Satoh S. 等,Region-dependent difference in the sleep-promoting potency of an adenosine A2A receptor agonist,European Journal of Neuroscience,第 11 卷第 5 期(1999 年 5 月):1587–1597。 ↩ ↩2
-
Radulovacki M. 等,Adenosine analogs and sleep in rats,The Journal of Pharmacology and Experimental Therapeutics,第 228 卷第 2 期(1984 年 2 月):268-274。 ↩
-
Strecker R.E. 等,Adenosinergic modulation of basal forebrain and preoptic/anterior hypothalamic neuronal activity,Behavioural Brain Research,第 115 卷第 2 期(2000 年 11 月):183-204。 ↩
-
Siegel J.M. 等,The Echidna Tachyglossus aculeatus Combines REM and Non-REM Aspects in a Single Sleep State,The Journal of Neuroscience,第 16 卷第 10 期(1996 年 5 月 15 日):3500-3506。 ↩
-
Plihal W., Born J., Effects of Early and Late Nocturnal Sleep on Declarative and Procedural Memory,Journal of Cognitive Neuroscience,第 9 卷第 4 期(1997 年 7 月):534-547。慢波睡眠剥夺主要影响陈述性记忆。 ↩ ↩2
-
Beersma D.G.M. 等,REM sleep deprivation during 5 hours leads to an immediate REM sleep rebound,Electroencephalography and Clinical Neurophysiology,第 76 卷第 2 期(1990 年 8 月):114-122。REM 反弹。 ↩
-
Crick F., Mitchison G., The function of dream sleep,Nature,第 304 卷(1983 年 7 月 14 日):111-114。反向学习理论。 ↩ ↩2 ↩3 ↩4
-
Stickgold R., Walker M.P., Sleep-Dependent Memory Consolidation and Reconsolidation,Sleep Medicine,第 8 卷第 4 期(2007 年 6 月):331–343。 ↩ ↩2
-
Hobson J.A., McCarley R.W., The brain as a dream state generator: an activation-synthesis hypothesis of the dream process,The American Journal of Psychiatry,第 134 卷第 12 期(1977 年 12 月 1 日):1335-1348。激活-合成理论。 ↩ ↩2 ↩3
-
Marshall L., Born J., The contribution of sleep to hippocampus-dependent memory consolidation,Trends in Cognitive Sciences,第 11 卷第 10 期(2007 年 10 月 1 日):442-450。 ↩ ↩2
-
Wilson M.A., McNaughton B.L., Reactivation of hippocampal ensemble memories during sleep,Science,第 265 卷第 5172 期(1994 年 7 月 29 日):676-679。慢波睡眠中海马体和新皮层网络重放放电模式。 ↩ ↩2 ↩3
-
Ficca G. 等,Morning recall of verbal material depends on prior sleep organization,Behavioural Brain Research,第 112 卷第 1-2 期(2000 年 7 月):159-163。 ↩ ↩2
-
Walker M.P. 等,Cognitive flexibility across the sleep–wake cycle: REM-sleep enhancement of anagram problem solving,Cognitive Brain Research,第 14 卷第 3 期(2002 年 11 月):317-324。 ↩ ↩2
-
Tononi G., Cirelli C., Sleep function and synaptic homeostasis,Sleep Medicine Reviews,第 10 卷第 1 期(2006 年 2 月):49-62。突触稳态假说。 ↩
-
Huber R. 等,Local sleep and learning,Nature,第 430 卷(2004 年 7 月 1 日):78-81。 ↩
-
Vyazovskiy V.V. 等,Cortical Firing and Sleep Homeostasis,Neuron,第 63 卷第 6 期(2009 年 9 月 24 日):865-878。 ↩
-
Massimini M. 等,Breakdown of Cortical Effective Connectivity During Sleep,Science,第 309 卷第 5744 期(2005 年 9 月 30 日):2228-2232。 ↩
-
Xu Q. 等, Day Napping and Short Night Sleeping Are Associated With Higher Risk of Diabetes in Older Adults,Diabetes Care,第 33 卷第 1 期(2010 年 1 月):78-83。 ↩ ↩2
-
Walker M.P., Cognitive consequences of sleep and sleep loss,Sleep Medicine,第 9 卷增刊 1(2008):29-34。 ↩ ↩2
-
Castellini M.A., Sleep in Aquatic Mammals. 载于 David W. Carley 和 Miodrag Radulovacki 编《Sleep-Related Breathing Disorders - Experimental Models and Therapeutic Potential》(纽约:Marcel Dekker, Inc.,2002),279-294。 ↩ ↩2
-
Fountain H., Study Gives Key Role to Sleep in Helping Brain Learn Anew,The New York Times。 ↩
-
Vyazovskiy V.V. 等,Cortical Firing and Sleep Homeostasis,Neuron,第 63 卷第 6 期(2009 年 9 月 24 日):865-878,doi: 10.1016/j.neuron.2009.08.024。 ↩
-
Tononi G., Cirelli C., Sleep function and synaptic homeostasis,Sleep Medicine Reviews,第 10 卷第 1 期(2006 年 2 月):49-62,doi: 10.1016/j.smrv.2005.05.002。 ↩
-
Squire L.R., Cohen N.J., Nadel L., “The medial temporal region and memory consolidation.” 载于 Weingartner H., Parker E.S. 编《Memory Consolidation: Psychobiology of Cognition》(新泽西:Lawrence Erlbaum Associates,1984),185-210。 ↩
-
McClelland J.L., McNaughton B.L., O’Reilly R.C., Why there are complementary learning systems in the hippocampus and neocortex,Psychological Review,第 102 卷第 3 期(1995 年 7 月):419-457。互补学习系统。 ↩ ↩2
-
Buzsáki G., The Hippocampo-Neocortical Dialogue,Cerebral cortex,第 6 卷第 2 期(1996 年 3-4 月):81-92。 ↩
-
Lee A.K., Wilson M.A., Memory of Sequential Experience in the Hippocampus during Slow Wave Sleep,Neuron,第 36 卷第 6 期(2002 年 12 月 19 日):1183-1194。 ↩
-
Wagner U. 等,Sleep inspires insight,Nature,第 427 卷(2004 年 1 月 22 日):352-355。睡眠激发洞见。 ↩ ↩2
-
Siegel J.M., The REM Sleep-Memory Consolidation Hypothesis,Science,第 294 卷第 5544 期(2001 年 11 月 2 日):1058-1063。 ↩
-
Stickgold R. 等,Sleep, Learning, and Dreams,Science,第 294 卷第 5544 期(2001 年 11 月 2 日):1052-1057。 ↩
-
Maquet P., The Role of Sleep in Learning and Memory,Science,第 294 卷第 5544 期(2001 年 11 月 2 日):1048-1052。 ↩
-
Vertes R.P., Siegel J.M., Time for the Sleep Community to Take a Critical Look at the Purported Role of Sleep in Memory Processing,Sleep,第 28 卷第 10 期(2005 年 10 月):1228-1229。 ↩
-
Walker M.P., The Role of Sleep in Cognition and Emotion,Annals of the New York Academy of Sciences,第 1156 卷(2009 年 3 月):168-197。 ↩
-
Tomita J. 等,No transcription-translation feedback in circadian rhythm of KaiC phosphorylation,Science,第 307 卷第 5707 期(2005 年 1 月 14 日):251–254。 ↩
-
Qin X. 等,Coupling of a Core Post-Translational Pacemaker to a Slave Transcription/Translation Feedback Loop in a Circadian System,PLoS Biology,第 8 卷第 6 期(2010 年 6 月 15 日):e1000394。 ↩